«Тест Плоские черви» - Тест: Плоские черви. А - ресничные, Б - сосальщики, В - ленточные. 6. Плоские черви имеют: 4. К какому классу плоских червей относится белая планария: А - лучевую симметрию, Б - двустороннюю, В - не имеют симметрию. 5. Чем покрыт наружный покровный слой у свободноживущих плоских червей: А - однослойные животные, Б - двухслойные животные, В - трехслойные животные.
«Кольчатые черви» - Есть боковые подвижные выросты тела – параподии. Представители типа. Класс Пиявки – медицинская пиявка, ложноконская пиявка, рыбья пиявка. Имеют кожно – мускульный мешок. Значение. Обладают: нервной, кровеносной, пищеварительной, выделительной, половой системами. Тип кольчатые черви. Среда обитания.
«Типы червей» - Длина от 80 мкм до 8 м; нитевидные или веретеновидные, реже бочонковидные или лимонообразные. Тип Круглые черви. Заднепроходного отверстия у П. ч. нет. Дробление яиц спиральное. Презентация по биологии ученицы 11 класса МОУ СОШ №1 Авралевой Любы. Органы выделения - метамерно расположенные парные нефридии.
«Тип Плоские черви» - Большинство имеет листообразную или лентовидную двусторонне-симметричную форму тела. Класс Ленточные (Cestoda). 8. Мускулатура: В наружном слое – кольцевые, во внутреннем – продольные мышцы. Фрагмент. Нервная система ортогонального типа, напоминает решетку. 6. Редия имеет ротовое отверстие и пищеварительную систему.
«Тип Кольчатые черви» - Строение концевой части тела. Выделительная система. Строение средней части тела. Имеют вторичную полость тела (целом). Тип кольчатые черви. Покров - кожно-мускульный мешок. 3. 1. Двусторонняя симметрия. Общие признаки кольчатых червей: Тип кольчатые черви черви. Нервная система. Вторичная полость тела кольчатых червей.
Всего в теме 9 презентаций
Класс олигохеты, или малощетинковые кольчецы (Oligochaeta)
Болынинство олигохет -обитатели почвы: более крупные из них хорошо всем известны под названием земляных или дождевых червей . Есть, однако, и пресноводные и даже морские олигохеты. Из общего числа (около 3 тыс.) описанных до сих пор видов примерно 400 - обитатели рек, озер, болот и других пресных водоемов, несколько десятков видов встречается в солоноватых водоемах и прибрежной зоне морей, все остальные - обитатели почвы (но некоторые из них, встречающиеся преимущественно по берегам водоемов, отчасти в воде, ведут земноводный образ жизни). В пределах СССР пока зарегистрировано немногим более 300 видов; цифра эта, однако, далеко не отражает действительного положения вещей, так как некоторые семейства, и в частности одно из наиболее богатых видами - энхитреиды , остаются у нас еще почти не изученными.
Среди олигохет есть и карлики и гиганты. Длина тела самых мелких пресноводных измеряется долями миллиметра, так что это в полном смысле слова микроскопические животные; а у наиболее крупных тропических земляных червей она превышает 2,5 м. Но это исключение. Большинство же водных олигохет имеет размеры от нескольких миллиметров до 10-15 см, а почвенных - от полусантиметра до 30-40 см.
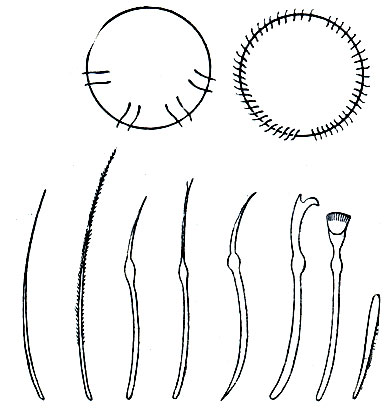
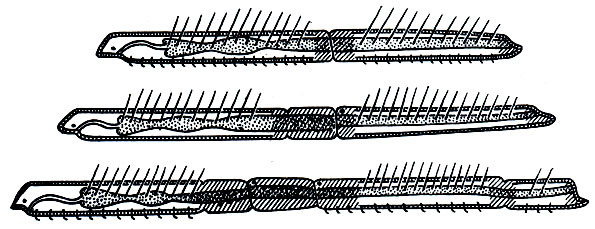
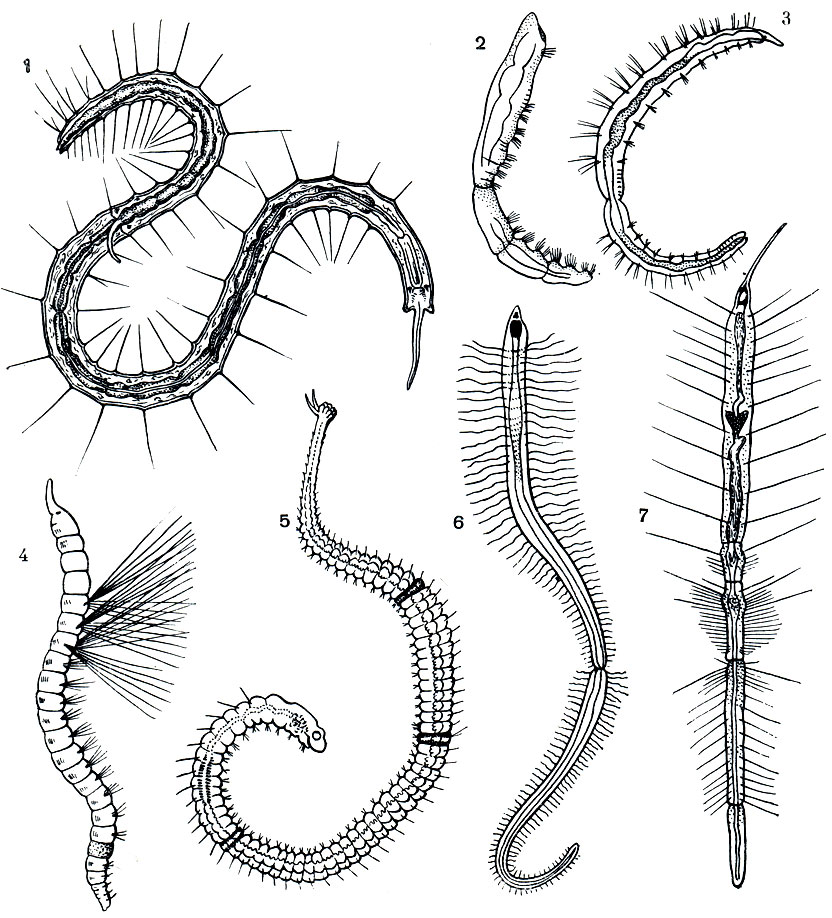
Важнейшей особенностью организации олигохет, как и полихет, является метамерия, т. е. правильная повторяемость органов вдоль оси тела животного. Внешне это выражается в том, что все червеобразное тело поделено перетяжками на отдельные участки - кольца (рис. 271), называемые сегментами или сомитами ("члениками"). Число сегментов тела у разных видов олигохет различно, от 5 - 6 до 500-600; в отличие от полихет они никогда не имеют парных выростов - парапо-дий, но несут по 4 пучка щетинок.
На переднем конце тела находится головная , или предротовая , лопасть ; у некоторых водных видов она бывает вытянута в более или менее длинный щупальцевидный придаток ("хоботок"), у немногих - почти исчезла, у большинства же представляет небольшой округленный выступ впереди рта. Далее следует первый, или ротовой, сегмент, никогда не несущий щетинок; на его нижней передней стороне находится рот. Все остальные сегменты, начиная со второго, как правило, снабжены четырьмя пучками щетинок каждый: двумя спинными (справа и слева) и двумя брюшными. У некоторых видов спинные пучки начинаются не вместе с брюшными, со второго сегмента, а несколько отступя от переднего конца тела - четвертого, пятого, шестого и даже двадцатого сегмента. Как исключение, имеются виды лишь с брюшными щетинками и даже вовсе без них.
В каждом пучке часто бывает по две более или менее одинаковых щетинки (например, у большинства дождевых червей). У тропических червей рода Pheretima число щетинок на каждом сегменте значительно больше - несколько десятков, а у некоторых видов до 150 - и расположены они не пучками, а в один ряд вокруг всего сегмента. У водных олигохет, как правило, в каждом пучке по нескольку щетинок, от 3-4 до 10-15, причем эти щетинки бывают различной формы: игловидные, крючковидные с простым или двузубчатым концом, веерные, простые и перистые волосовидные и др. (рис. 272).
Стенка тела олигохет состоит из пяти основных слоев: тонкой неклеточной кутикулы, выделяемой кожей, кожного эпителия, состоящего из одного слоя клеток, двух слоев - мускулатуры наружного кольцевого и внутреннего продольного и, наконец, внутреннего слоя целомического эпителия, ограничивающего вторичную полость тела, или целом, в которой располагаются внутренние органы. Эта полость, заполненная более или менее прозрачной жидкостью, содержащей взвешенные в ней клетки, не тянется непрерывно вдоль всего тела, а разделена тонкими мускульными перегородками - диссепиментами - на отдельные отсеки, соответствующие наружной сегментации. Таким образом, в каждом сегменте находится свой обособленный участок полости тела, а в нем - пара своих выделительных органов - нефридиев, один справа, другой слева (рис. 273). Имеется замкнутая кровеносная система: продольные сосуды идут вдоль всего тела, связывающие их кольцевые сосуды имеются в каждом сегменте. Пищеварительный канал проходит вдоль всего тела от ротового отверстия до анального; обычно он подразделяется на отделы: ротовая полость, глотка, пищевод, иногда зоб, один или несколько желудков, средняя кишка (самая длинная часть), задняя кишка (рис. 274).
В каждом сегменте, в брюшной его части, находится двойной нервный узел (состоящий из сближенных и почти слившихся правого и левого) с отходящими от него нервами. Узлы всех сегментов соединяются продольными нервными тяжами - коннективами - в единую брюшную нервную цепочку. В головной лопасти или несколько позади нее, над глоткой, находится головной мозг (надглоточный ганглий), соединенный с первым узлом брюшной цепочки - подглоточным - двумя тяжами, охватывающими глотку,- окологлоточными коннективами . Глаз у большинства олигохет нет, они имеются только у части видов одного семейства водных олигохет - наидид ; однако, как правило, олигохеты реагируют на освещение, что связано с наличием в коже, особенно на головном конце, особых светочувствительных клеток фоторецепторов . Дыхание у подавляющего большинства кожное, но у некоторых пресноводных олигохет имеются жабры.
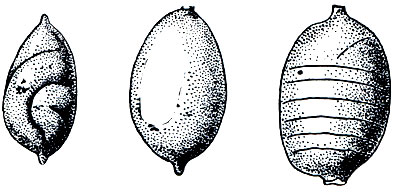
Все олигохеты гермафродиты, т. е. у каждого взрослого червя имеются одновременно и мужская и женская половые системы, строение которых довольно сложно (рис. 275). Оплодотворению предшествует спаривание, во время которого сперматозоиды каждого из двух червей переходят в семеприемники другого (рис. 276). После этого черви расходятся. Затем у каждого из них на пояске, представляющем железистое утолщение кожи нескольких определенных сегментов (у разных олигохет различных, но всегда ближе к головному концу), выделяется кокон, первоначально имеющий форму бочонка (или муфты), открытого с обоих концов. Образовавшийся кокон сокращениями мускулатуры тела сдвигается постепенно вперед и наконец сбрасывается "через голову", после чего концы его смыкаются и он принимает форму лимона (рис. 277). Но прежде чем кокон будет сброшен, пока он медленно скользит от пояска к головному концу, в него откладываются яйца (когда он продвигается над отверстиями яйцеводов), а затем из семеприемников поступают сперматозоиды, полученные при спаривании от другого червя. Таким образом, оплодотворение яиц происходит не в теле червя и не во внешней среде, а в коконе, внутри которого происходит и дальнейшее развитие. Из кокона выходят маленькие червячки, в общем уже похожие на взрослых (рис. 278). У некоторых олигохет описано самооплодотворение, а другие способны к партеногенезу, т. е. девственному, без оплодотворения, развитию отложенных яиц.
Кроме полового, у олигохет наблюдается и бесполое, вегетативное, размножение, но оно свойственно представителям только некоторых семейств водных червей. Бесполое размножение может происходить либо путем архитомии , либо путем паратомии . При архитомии тело червя самопроизвольно (иногда в результате внешнего раздражения) распадается на два или большее количество кусков - фрагментов, после чего у каждого куска развиваются недостающие части и он постепенно превращается в целого червя.
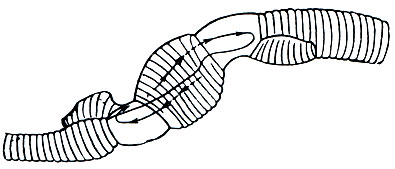
При паратомии (рис. 279) на теле червя, на некотором определенном расстоянии от головного конца намечается перетяжка, подразделяющая его на две части; после этого задняя часть, не отделяясь сначала от передней, образует свой головной конец, а передняя - свой задний. Теперь два образовавшихся таким образом червя могут разделиться окончательно, но часто, прежде чем это произойдет, каждый из них опять начинает делиться. Таким образом возникают "цепочки", состоящие из четырех-пяти, а иногда и большего числа особей.
Деление класса олигохет на отряды еще окончательно не установилось; мы рассмотрим последовательно важнейшие семейства.
Семейство Эолосомы (Aeolosomatidae) включает около полутора десятков видов самых мелких пресноводных олигохет, длина тела которых редко превышает 2- Змм, а у некоторых даже меньше 1 мм. Поэтому, хотя они встречаются довольно часто в болотах, канавах и прибрежных зарослях прудов, озер и рек, обнаружить их не так легко, а хорошо рассмотреть можно только под микроскопом. Зато если среди водорослей и простейших в капле воды удастся заметить одно из этих изящных животных, например гемприхову эолосому (Aeolosoma hemprichi), одну из самых обычных у нас, понаблюдать за ним стоит. Маленький прозрачный червячок сразу узнается по характерным оранжевым кожным железкам, которые разбросаны по всей поверхности тела в виде округлых пятнышек (рис. 280). У других видов эолосом железки окрашены иначе - в желтоватый, зеленый или голубоватый цвета, а могут быть и бесцветными. Число сегментов их тела невелико - от 7 до 20, сегментация выражена не резко. На каждом сегменте тела имеется по четыре пучка тонких волосовидных щетинок, а на нижней стороне головной лопасти и около рта - многочисленные реснички, благодаря которым эти черви могут не только ползать, но и плавать; для них характерно скользящее движение. Пищей им служат микроскопические одноклеточные водоросли и бактерии, мельчайшие частицы разлагающихся органических веществ, поэтому особенно благоприятные условия для размножения большинство из них находит в умеренно загрязненных водоемах, в местах скопления гниющих растений у берегов, в болотах и т. п. Нередко эолосомы размножаются в большом количестве в банках, где разводят в лаборатории инфузорий и других простейших, а также иногда в аквариумах с рыбами, для которых они, конечно, никакой опасности не представляют. Половое размножение у эолосом наблюдается редко, обычно они размножаются вегетативно, путем паратомии (см. выше), причем образуются цепочки из двух-пяти особей.
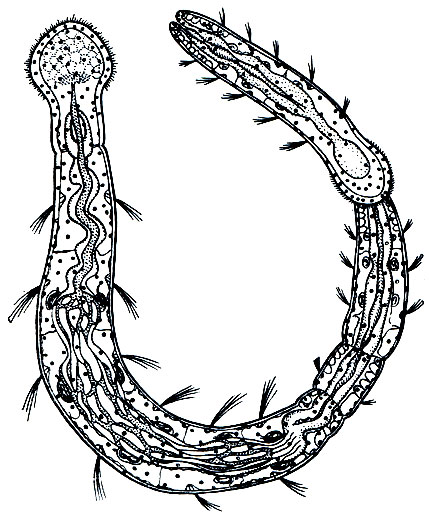
Семейство Наидиды , или Водяные змейки (Naididae), гораздо богаче видами, которых насчитывается свыше сотни. Все они - водные животные, причем большинство является обитателями пресных водоемов и лишь немногие встречаются в солоноватых водоемах и в прибрежной зоне морей. Хотя среди наидид имеются и очень мелкие, длиной всего в 1 - 2 мм, большинство из них крупнее, 10-20 мм, и хорошо заметны простым глазом, а самые крупные достигают длины 35-50 мм. Сегментация выражена хорошо, число сегментов у большинства в пределах от 10-15 до 30-40, но есть виды, имеющие до 100 сегментов. По форме тела, внешнему виду, окраске наидиды очень разнообразны. Головная лопасть у большинства округлая или удлиненная, но у некоторых вытянута в более или менее длинный щупальцевидный придаток ("хоботок"), а у других укорочена и почти совсем не развита. Наидиды - единственное семейство олигохет, у большинства представителей которого имеются глаза. Щетинки, как правило, расположены на каждом сегменте четырьмя пучками (2 спинных и 2 брюшных), причем форма щетинок очень разнообразна: в брюшных пучках чаще всего имеются вильчатые крючковидные, а в спинных - игловидные и более длинные волосовидные; но есть виды, лишенные волосовидных щетинок, а есть и такие, у которых вообще отсутствуют спинные пучки, так что щетинки имеются только на брюшной стороне. Спинные щетинки у большинства, в том числе у видов рода Nais, начинаются с шестого сегмента, но у пристин (род Pristina) - со второго, а у некоторых других - с третьего, четвертого, пятого или позади шестого.
Некоторые виды наидид имеют жабры, расположенные у одних на заднем конце, а у других вдоль большей части тела, по бокам.
У наидид тоже имеются две формы размножения: половое, обычно приуроченное к определенным сезонам, и вегетативное, путем паратомии, которое преобладает; поэтому чаще встречаются не одиночные черви, а "цепочки", состоящие из двух - пяти особей.
Наидиды встречаются во всех странах мира; в СССР обнаружено свыше 50 видов.
К числу наиболее обычных в наших водоемах принадлежат многие виды рода наид (Nais). Редко случается, чтобы их не удалось обнаружить при рассматривании в банке воды, зачерпнутой вместе с водными растениями у берега пруда, озера, речки или в болоте. Большинство видов этого рода - нежные беловатые прозрачные червячки длиной около 10-15 мм, с торчащими в стороны пучками тонких щетинок и парой темных глаз на округлоудлиненной головной лопасти (рис. 281). Они оживленно ползают, поочередно вытягивая и сокращая свое тело, нередко обвиваясь вокруг нитчатых водорослей; многие виды этого рода хорошо плавают. Наиды питаются растительной пищей и мельчайшими частицами разлагающихся остатков других организмов; в их кишечнике постоянно можно видеть одноклеточные зеленые и диатомовые водоросли, а иногда и некоторых простейших животных. По-видимому, немалую роль в их питании играют и бактерии. В аквариумах многие наиды хорошо живут и энергично размножаются при подкормке дрожжевой взвесью. Отдельные виды этого рода различаются с трудом, поэтому мы не приводим здесь их описания; отметим только, что есть среди них виды, встречающиеся преимущественно в реках (например, Nais behningi), есть связанные преимущественно с холодными ручьями и родниками (Nais elingius), большинство же встречается в прибрежной зоне как различных стоячих водоемов, так и рек (Nais variabilis, N. pseudoobtusa и др.).
Как уже упоминалось, у большинства олигохет дыхание кожное, но у некоторых имеются специальные органы дыхания - жабры. Среди семейства наидид обладателями жабр являются виды широко распространенных родов Dero и Aulophorus (рис. 281), а также сравнительно редкий Branchiodrilus hortensis, найденный в СССР только в бассейне реки Амура и в озере Ханка (рис. 281).
Dero и Aulophorus - обитатели дна водоемов. Передним концом они зарываются в ил (которым питаются), а задний выставляют над поверхностью дна. Хвостовой конец у них расширен в виде диска, на котором сидят парные листовидные жабры. Эти жаберные листочки густо покрыты ресничками, благодаря работе которых свежая вода все время омывает жабры.
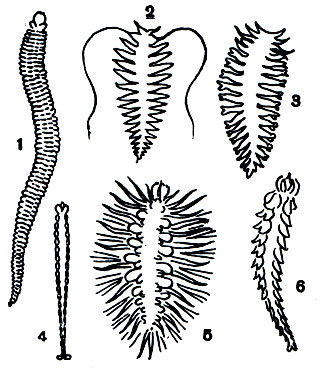
Виды родов Dero и Aulophorus имеют красноватую окраску, зависящую от наличия в их крови гемоглобина, что в свою очередь связано с необходимостью лучше использовать кислород, которого в придонных слоях воды может быть очень мало. Глаз эти донные виды не имеют. Некоторые из них хорошо плавают.
Некоторые мелкие наидиды, особенно виды рода Pristina, лишенные глаз (рис. 281), нередко в большом количестве размножаются в аквариумах с рыбами. У всех пристин спинные щетинки начинаются со второго сегмента, как и брюшные; у некоторых головная лопасть вытянута в небольшой хоботок. Никакой опасности для рыб они не представляют, скорее даже наоборот: питаясь иловыми частицами, бактериями, одноклеточными водорослями, они являются хорошими санитарами, а кроме того, ими самими могут питаться мальки рыб.
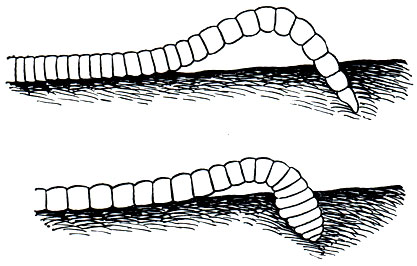
Семейство Трубочники , или Тубифициды (Tubificidae), включает около 130 видов олигохет, подавляющее большинство которых обитает на дне пресных водоемов и лишь немногие встречаются в прибрежной зоне морей. В СССР найдено около 70 видов этого семейства. Трубочники крупнее наидид; их тонкое нитевидное тело (рис. 282) достигает у некоторых видов длины 100-180 мм, большинство же имеет длину 30-60 мм, а самые мелкие - всего 5-10 мм. Все они лишены глаз. Тело состоит из довольно большого количества сегментов, обычно 60-100, у некоторых - свыше 200. Щетинки расположены четырьмя пучками на каждом сегменте, причем в брюшных пучках имеются только простые, крючковидные или вильчатые, а в спинных у многих видов еще и волосовидные. Трубочники - типичные донные животные, особенно многочисленные на илистых грунтах; нередко они образуют массовые скопления, выглядящие как красноватые "подушки" на дне и состоящие из тысяч особей. Некоторые виды выдерживают очень сильное загрязнение и могут жить при минимальном количестве растворенного в воде кислорода, перенося в течение многих суток даже полное его отсутствие. Все трубочники имеют красную или розоватую окраску, обусловленную наличием в их крови гемоглобина; именно наличие гемоглобина, а также свойственная им способность производить особые дыхательные движения, вызывающие приток к их телу свежей воды (рис. 283), и позволили им заселить дно илистых водоемов. Следует помнить, что илы благодаря происходящим в них процессам разложения органических остатков поглощают кислород из придонных слоев воды, снижая здесь его количество до минимальных величин. Передним концом трубочники обычно погружены в ил, а задний выставляют над поверхностью дна, производя им колебательные (дыхательные) движения (рис. 283). Именно в коже заднего конца тела особенно густо разветвлены кровеносные сосуды, так что хвост червя является в известном смысле органом дыхания. Чем меньше в окружающей воде кислорода, тем больше высовываются хвосты червей из ила и тем энергичнее они производят дыхательные движения. Наоборот, при высоком содержании кислорода черви почти полностью погружаются в ил. Вокруг высовывающейся из грунта части тела многие виды трубочников образуют из склеенных слизью иловых частиц короткую гибкую трубку - отсюда и их название. Потревоженные трубочники мгновенно скрываются в иле.
Трубочники - типичные грунтоеды. Погружаясь передним концом в грунт дна водоема на глубину до 50-100 мм, они все время заглатывают ил с песком, пропуская через кишечник за сутки количество материала, во много раз превышающее вес их собственного тела. При этом, естественно, происходит минерализация пропускаемого через кишечник грунта, так как входящие в его состав органические вещества частично используются червями для питания. Характерно, что, чем беднее грунт органическим веществом, тем больше его приходится заглатывать и пропускать через свой кишечник червям.
Если учесть, что в сильно загрязняемых водоемах, например реках и прудах в черте городов, количество трубочников может достигать более ста тысяч на 1 м 2 дна, легко представить себе масштабы их деятельности и ее значение в процессах биологического самоочищения этих водоемов. Хотя таких высоких цифр количество трубочников достигает только в сильно загрязненных, богатых органическими веществами местах, их все же достаточно и на дне более или менее чистых рек, озер и водохранилищ: сотни и тысячи на каждый квадратный метр. Это значит, что они играют немалую роль в питании рыб, тем более что тело их, богатое ценными питательными веществами, очень легко переваривается в кишечнике рыб. Трубочники являются очень удобным и хорошим кормом и для аквариумных рыб.
Размножение у них только половое. После спаривания черви откладывают коконы, содержащие по нескольку яиц каждый. Если рассматривать трубочников невооруженным глазом, то различные их виды трудно отличить друг от друга; только изучив под микроскопом особенности строения спинных и брюшных щетинок, можно в них разобраться, а для точного определения вида часто и этого бывает недостаточно, приходится изучать строение половой системы. Поэтому мы не приводим здесь описания различных видов (для этого существуют определители, например, О. В. Чекановская, "Водные малощетинковые черви фауны Советского Союза"), укажем только, что наиболее часто встречающимися являются виды рода Tubifex, имеющие в спинных пучках волосовидные и крючковидные, или веерные, щетинки, например самый обычный из них и особенно стойкий к загрязнению трубочник обыкновенный (Tubifex tubifex).
Виды рода Limnodrilus лишены волосовидных щетинок; из них чаще других, нередко вместе с Т. tubifex, встречается лимнодрил Гоффмейстера (Limnodrilus hoffmeisteri), у которого все щетинки, и спинные и брюшные, имеют по 2 загнутых одинаковых зубца на конце. Назовем еще самого крупного из наших лимнодрилов - невского (L. newaensis). Он впервые был найден в реке Неве, но оказался широко распространенным в реках, водохранилищах и некоторых озерах Европейской части СССР.Это довольно толстый, до 2 мм, червь, длиной до 90 мм, обычно темной серовато-красной окраски; щетинки у него с короткими зубцами, причем верхний короче нижнего.
В прибрежной полосе морей, в частности Белого и Баренцева, при солености до 20‰, обитает не встречающийся в пресных водоемах Clitellio arenarius, как и лимнодрилы, лишенный волосовидных щетинок.
Есть среди трубочников и обладающие наружными жабрами. Это довольно редкий вид- бранхиура (Branchiura sowerbyi, рис. 284). Водится бранхиура в Индии, Индонезии, Китае и Японии, была найдена во Франции, а у нас обнаружена в бассейне реки Амура и в Закавказье. Интересно, что этот вид много раз находили в тепловодных бассейнах ботанических садов, в которых выращивают знаменитую викторию (Victoria regia). Впервые он был найден именно в таком бассейне в Лондоне и описан; позже он был найден в бассейне оранжереи ботанического сада Московского университета.
Семейство Энхитреиды (Enchytraeidae) содержит как водных, так и сухопутных (почвенных) червей, причем последних больше, а общее количество видов этого семейства достигает почти 400. Энхитреиды - небольшие или мелкие, малопрозрачные черви, в большинстве случаев беловатые, реже желтоватые или розоватые. Длина их от 2-З мм до 40-45 мм, большинство видов от 10 до 25 мм. Сегментация выражена хорошо, число сегментов, как правило, в пределах от 25 до 50, у некоторых больше, до 70-80. Глаз нет.
Щетинки расположены четырьмя пучками на каждом сегменте, начиная со второго; все щетинки более или менее одинаковые, игловидные или палочковидные, с простым, не вильчатым (за одним исключением) кончиком. Размножение только половое; у некоторых наблюдается своеобразная форма бесполого размножения, при котором червь распадается на две или большее число частей, восстанавливающих затем путем регенерации недостающие головные или хвостовые отделы (так называемая архитомия). При благоприятных условиях некоторые виды почвенных и литоральных энхитреид размножаются в огромном количестве, до десятков тысяч на 1 м 2 (а в очень богатых перегно.ем почвах даже до 150-200 тыс.), и играют очень существенную роль в разложении органических остатков и почвообразовании. Благодаря тому что их легко разводить в ящиках или других сосудах с землей (размножаются они очень быстро), некоторые виды, главным образом белый энхитрей (Enchytraeus albidus), стали использоваться как хороший корм для рыб, притом не только аквариумных, но и в более крупных масштабах, на рыбоводных заводах. Поскольку эти энхитреиды разводились аквариумистами часто в цветочных горшках с землей, они получили название "горшечных червей". На морских побережьях, в частности на Белом море, под выброшенными волнами гниющими водорослями некоторые энхитреиды, особенно Enchytraeus albidus и Lumbricillus lineatus, образуют большие скопления, в которых черви буквально кишат.
Распространены энхитреиды очень широко, преимущественно в северном полушарии, в умеренной и субарктической зонах; на север они проникают дальше других олигохет и, в частности, известны со Шпицбергена, Новой Земли, Новосибирских островов и Гренландии. Встречаются и высоко в горах, до высоты 3-4 км над уровнем моря, найдены и в пещерах.
Известно, что некоторые виды этого семейства выдерживают продолжительное воздействие температуры ниже нуля. Так, ряд ученых наблюдал их оживание при оттаивании мерзлого грунта или кусков льда, в который они вмерзли. Американский зоолог М у р рассказывает, что он наблюдал один из видов этих червей (Mesenchytraeus solifugus) на снегах горных ледников Аляски; в начале лета, когда снег был плотным, они вяло ползали по его поверхности вечером и ночью, но в середине дня, когда солнце светило ярко, они зарывались вглубь; позже, когда снег стал подтаивать и пропитался водой, они были уже более активны и нередко их можно было наблюдать в образовавшихся на поверхности лужицах. Подобные же наблюдения над этим и другими видами были сделаны и на других ледниках. Интересно, что многие виды энхитреид, живущие постоянно на снегах, имеют, как и встречающиеся там же мелкие насекомые, черную или темную красно-коричневую окраску; возможно, что это имеет значение для поглощения тепловых лучей и как защита от ультрафиолетового излучения. Пищей им могут служить одноклеточные красные водоросли, часто в несметном количестве размножающиеся на горных снегах, а также, возможно, пыльца хвойных.
Семейство Гаплотаксиды (Нарlotaxidae) включает всего около 20 видов водных олигохет. Из этого небольшого семейства мы упомянем один широкораспространенный вид - Haplotaxis gordioides. Это очень длинный, до 300 мм, и тонкий червь, тело которого состоит из 200-480 сегментов (рис. 285); каждый сегмент несет всего по 4 или даже только по 2 (брюшные) довольно крупные простые щетинки. Встречается этот червь, однако, нечасто и только местами; держится преимущественно в водоемах с холодной водой - колодцах, источниках, ручьях, реках и совершенно отсутствует в сильно прогреваемых прудах и т. п.; в больших озерах отмечен до глубины свыше 300 м. Иногда попадается и не в воде, а в глинистом грунте по берегам ручьев и других водоемов. Чаще его находят в весенние месяцы. Есть указание, что, попав в аквариум, эти черви могут повреждать корни растений, например валлиснерии. В СССР Haplotaxis gordioides найден в озере Байкал, где водится и другой представитель этого рода-Haplotaxis ascaridoides (нигде, кроме Байкала, не найденный), в Телецком озере на Алтае (где, кроме него, водится другой представитель этого семейства - Pelodrilus ignatovi), а также в озерах Камчатки, реках и озерах Кавказа, Средней Азии.
Семейство Люмбрикулиды (Lumbriculidae) включает свыше 60 видов. Они имеют только крючковидные щетинки, одно- или двухзубчатые, по 4 пары на каждом сегменте, начиная со второго. Головная лопасть у большинства округленная, у некоторых вытянута в небольшой хоботок, глаз нет. Окраска обычно красноватая или серовато-красная, иногда с фиолетовым оттенком, иногда, преимущественно на переднем конце, с зеленоватой или черноватой пигментацией. Тело нередко полупрозрачное, его поверхность у некоторых видов при сильном освещении отливает радужными тонами (иризирует). Люмбрикулиды - обитатели пресных вод. Большинство из них имеет средние размеры от 20-30 мм до 80-100 мм при толщине 1 - 2 мм, но наиболее крупные виды достигают длины свыше 180 мм и толщины до 4,5 мм. Число сегментов у большинства видов около 80-150, но у некоторых больше 200.

Самым обычным представителем семейства является Lumbriculus variegatus, встречающийся почти во всех водоемах и очень широко распространенный. Особенно часто он попадается в небольших заболоченных водоемах, лужах, канавах. Его длина - 40-90 мм, окраска темная, красноватая. Легче всего узнать люмбри-кулюса по характерным движениям: будучи потревожен, он начинает быстро и резко извиваться и уплывает в сторону. Особенно ярко эта реакция бывает выражена в скоплениях червей: такой клубок, если его потревожить, буквально "взрывается", и все черви бросаются в разные стороны. Однако сколько-нибудь долго люмбрикулюсы плыть не могут и вскоре снова опускаются на дно. Другой особенностью этого вида является то, что он почти утратил способность к половому размножению; найти половозрелых червей почти никому не удавалось. Зато он приобрел способность очень быстро размножаться бесполым путем, распадаясь на отдельные куски (размножение фрагментацией, или архитомией), которые затем восстанавливают головные и хвостовые концы путем регенерации. Люмбрикулюса почти невозможно схватить пинцетом не повредив: он сейчас же распадается на 2 или несколько кусков. Способность к регенерации у этого вида очень велика, и червь может восстановиться даже из небольшого куска.
Другой широко распространенный, но более редкий вид этого семейства- Rhyn-chelmis limosella - значительно крупнее: 80-150 мм длиной, 2-3 мм толщиной. Встречается преимущественно в холодных или сильно затененных водоемах, обладает "хоботком".
Оба вида послужили объектами многих специальных исследований по изучению закономерностей процесса регенерации.
Но самой, может быть, замечательной особенностью этого семейства является то, что больше половины всех его видов (около 40) водится только в озере Байкал и нигде больше на земном шаре не встречается! Люмбрикулиды заселяют Байкал от прибрежной полосы до глубин свыше 1 км, причем каждой зоне свойственны свои виды. Они очень разнообразны и по размерам и по внешнему виду, некоторые обладают характерной окраской и сильно иризируют. Это обстоятельство, т. е. наличие в Байкале большого количества эндемичных (только в нем встречающихся) видов люмбрикулид и их своеобразие, говорит о большой древности Байкала именно как пресноводного озера.
Семейство Настоящие дождевые черви , или Люмбрициды (Lumbricidae). Всем хорошо известны земляные, или дождевые, черви. Иногда в сырую погоду, особенно после дождей (отсюда и название), они в большом количестве появляются на поверхности, ползают по дорожкам садов и парков, лежат на дне лужиц дождевой, а весной талой воды. Не следует, однако, думать, что название "дождевые черви" обозначает какое-то одно определенное семейство олигохет; оно применяется ко всем более или менее крупным представителям пяти различных семейств этого класса, обитающим в почве, т, е. является сборным. На первый взгляд, они действительно до некоторой степени сходны между собой; зоологи различают эти семейства главным образом по особенностям строения внутренних органов, в частности органов размножения, по расположению пояска, щетинок и по другим признакам. В учебниках зоологии в качестве примера для класса олигохет приводится "обыкновенный дождевой червь" - Lumbricus terrestris. Этот довольно крупный червь, до 30 см длиной и до 1 см толщиной, принадлежит к наиболее полно изученному семейству настоящих дождевых червей , или люмбрицид (Lumbricidae), включающему около 200 видов, самые мелкие из которых имеют всего 2-3 см в длину и около 1 мм в толщину, а самые крупные достигают полуметровой длины при толщине до 1,5-2 см.Размножение у них только половое.
Упомянутый нами Lumbricus terrestris, носящий местами народные названия "большой выползок" или "большой красный червь", легко отличается от других дождевых червей своими крупными размерами, уплощенным и расширенным ("лопатообразным") хвостовым концом и окраской: передняя треть его тела со спинной стороны довольно темного, тусклого красно-фиолетового цвета, а остальное тело светлее, грязно-розоватого оттенка, с более темной полосой, тянущейся вдоль середины спины почти до самого конца тела. На солнце передний конец этого червя отливает радужными тонами (иризирует), что связано с особенностями строения кутикулы. Щетинки у него расположены на сегментах четырьмя парами, а поясок - с 31-го или 32-го по 37-й сегмент.
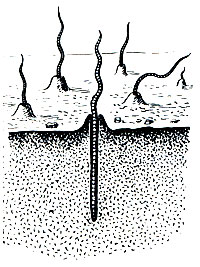
Этот вид распространен по всей Европе, от Атлантического побережья до Урала; в Сибири, Казахстане, Средней Азии, на Кавказе не найден, хотя не исключено, что может быть местами обнаружен, как завезенный, в городских садах и парках; известен он и в Северной Америке. В земле он роет глубокие ходы ("норы") уходящие от поверхности на 2 м и больше. В теплую и сырую погоду, по ночам * эти черви вылезают из своих норок, но не совсем, а цепляясь хвостом за отверстие, так что в случае опасности могут быстро скрыться. Вытягиваясь, они обшаривают окружающее пространство, захватывают своим ртом (при этом слегка выворачивается наружу, а затем втягивается обратно мускулистая глотка) сырые опавшие листья, полусгнившие травинки и т. п. и затаскивают их в свои норы. Но черви заглатывают и землю, которая поэтому всегда имеется в их кишечнике. Органические вещества почвы, перегной, используются в качестве пищи, песчинки способствуют измельчению и перетиранию проглоченных кусочков листьев, наконец, частично почва заглатывается при прорывании ходов, хотя главная работа при этом проделывается мускулатурой стенки тела червя. Когда червь зарывается в землю с поверхности (рис. 286) или когда прокладывает новый ход в толще почвы, он действует передним концом как клином, попеременно то сужая и вытягивая его, то вздувая и сокращая, и таким образом расталкивает частицы почвы в стороны. Большую роль при этом играют щетинки, цепляющиеся за стенки хода и не дающие возможности телу червя соскальзывать назад при его сокращении. По поверхности земли, не имея опоры со всех сторон, черви передвигаются сравнительно медленно, но внутри своих ходов - довольно быстро: если перевернуть камень или другой предмет, под которым скопились черви, и попытаться схватить их, они почти мгновенно скрываются в норки.
* (Дождевые черви вообще очень чувствительны к свету. При внезапном освещении дневным светом, например при переворачивании камней, под которыми они скопились, или ночью при свете фонаря они быстро скрываются в норки. Однако на очень слабый свет они реагируют положительно. К красному свету практически нечувствительны. )
Переработанную ими, прошедшую через кишечник землю черви выбрасывают на поверхность над отверстиями ходов в виде характерных кучек - выбросов , или копролитов . Появление таких кучек на оголенной земле весной свидетельствует о начале деятельности червей. Подсчитано, что за сутки каждый червь пропускает через свой кишечник количество земли, примерно равное весу его тела (а средний вес Lumbricus terrestris - 4-5 г). Зимуют эти черви в нижней части своих ходов, ниже глубины промерзания почвы; при высыхании почвы во время летней засухи они также спускаются в глубокие ее горизонты. Продолжительность жизни этого вида точно неизвестна, но, по некоторым наблюдениям, не меньше 5-6 лет.
К роду Lumbricus принадлежит 10 видов. Самым обычным у нас является малый красный червь , или малый выползок (Lumbricus rubellus). Он примерно вдвое короче и тоньше предыдущего, но окрашен ярче, вишнево-красного цвета. Задний конец его тела тоже уплощен и расширен, а поясок у него обычно расположен на сегментах с 27-го по 32-й. В средней полосе это, пожалуй, второй по встречаемости вид. Таких глубоких ходов, как большой выползок, он не делает; обычно держится в самых поверхностных горизонтах почвы, а в лесах - в сыроватой подстилке, среди полугнилых листьев.
Но самым распространенным и часто встречающимся у нас видом является серая аллолобофора (Allolobophora caliginosa), называемая иногда пашенным червем, так как на пашнях и полях попадается чаще других; найти ее можно также и в лесах, на огородах и на побережьях водоемов. Это средних размеров, до 15-16 см, длиной, сероватый червь, совершенно лишенный пурпурной пигментации, с пояском, расположенным обычно на сегментах с 27-го по 34-й. Задний конец тела не уплощен. Как у других видов этого рода (а их насчитывается около 50), щетинки на каждом сегменте сближены попарно. В отличие от выползков аллолобофора держится обычно в толще почвы, на глубине 5-15 см, где и питается главным образом почвенным перегноем. На поверхность ее могут выгнать только сильные дожди. При благоприятных условиях размножается в огромном количестве: с площади в 1 м 2 удается иногда выкопать до 400-500 экземпляров. Во время летней засухи серая аллолобофора уходит на глубину 40-60 см, свертывается там клубком, иногда буквально завязываясь узлом, выделяет слизь и с ее помощью строит вокруг себя из почвенных частиц довольно прочную защитную капсулу, впадая затем в неактивное состояние, причем процессы развития приостанавливаются. Такое состояние, диапауза, может продолжаться больше двух месяцев. Благодаря этой способности аллолобофоры более устойчивы к засухе, чем выползки, и могут заселять более сухие, открытые места.
К этому роду принадлежит и один из самых крупных наших червей - алтайская гигантская аллолобофора (Allolobophora magnifica), носящая у местных жителей названия "щура" и "салазана". Длина заспиртованных червей этого вида достигает 25-30 еле при толщине около 1,5 см; живые черви могут растянуться до 40 см и больше. Их ходы, до 2 см диаметром, проникают в почву на глубину 4-5 м. Местами вся почва бывает пронизана такими ходами, а над отверстиями обитаемых норок образуются кучки выбросов, достигающие 8 см в высоту и 10 см в поперечнике. С сельскохозяйственной точки зрения воздействрге на почву этих червей (как и других видов) положительно, но строители смотрят на это дело часто иначе, так как сильное рыхление червями земли около фундаментов доставляет им лишние хлопоты. Пока этот вид известен только из районов Северо-Западного Алтая.
Род Дендробена (Dendrobaena) характеризуется широко расставленными (не сближенными попарно) щетинками. Название рода указывает на обитание его представителей в лесах; описанный уже давно вид этого рода - небольшой, очень подвижный, темный, красно-фиолетовый червь восьмигранная дендробена (Dendrobaena octaedra)-настолько характерен для лесной зоны, что нередко его называют "таежным червем". Держится он главным образом под сырыми опавшими листьями, в лесной подстилке, и распространен широко - от Атлантического океана до Тихого, встречаясь на далеком Севере, в Заполярье и в горах Крыма и Кавказа.
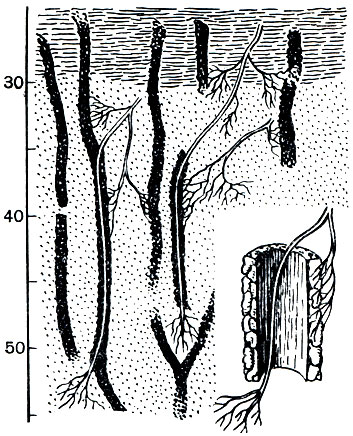
Однако далеко не все виды этого рода, а их насчитывается свыше 40, являются обитателями лесов. Мы упомянем еще только один из них - мариупольскую дендробену (Dendrobaena mariupolienis), распространенную в Приазовье, Крыму и на Кавказе. Это очень крупный червь, до 35 см длиной, довольно толстый и сильный, темной, черновато-бурой окраски; поясок у него захватывает с 25-го по 32-й сегмент. По наблюдениям Г. Н. Высоцкого, мариупольская дендробена довольно многочисленна в почвах целинных степей. Замечательно, что ходы этого червя удалось проследить на глубине 8 м; часто по ним проникают в нижние горизонты почвы корни растений (рис. 289).
Одним из наиболее ярко окрашенных червей является навозный (Eisenia foetida), обладающий характерным неприятным запахом, что отражено и в его названии - foetida, что значит "дурно пахнущая", "вонючая". Это сравнительно небольшой червь, длиной 6-10 см, с попарно сближенными щетинками. Цвет его темно-красный или красно-коричневый, но окраска не сплошная, а кольчатая, так как бороздки, разделяющие сегменты, светлые, что особенно хорошо видно, когда червь сильно растянется. Навозный червь - очень широко распространенный вид; на севере и в средней полосе он встречается почти исключительно недалеко от жилья человека, в скоплениях навоза, гнилой соломы, в парниках и т. п. (нередко массами, до 1000 и более на 1 м 2). Но уже в лесостепной полосе, а также в Крыму и на Кавказе он становится довольно обычным и в лесах, далеких от жилья, где его можно найти в старых, трухлявых пнях, дуплах, скоплениях прелых листьев и по берегам речек и ручьев. При раздражении навозный червь выделяет довольно сильно пахнущую желтую жидкость.
К этому же роду относится и норденшельдов червь , или эйзения Норденшельда (Eisenia nordenskioldi), названная так в честь известного шведского путешественника, исследователя полярных стран Норденшельда , собравшего большие зоологические коллекции на севере Сибири. Этот вид распространен по всей Сибири, встречается на Алтае и в Маньчжурии, а также в восточной части Европейской территории СССР, доходя до Крыма. По окраске и размерам он очень изменчив; типична для него окраска, похожая на таковую у Lumbricus rubellus: довольно темная, вишнево-красная, причем задний конец несколько светлее; характерны два светлых пятна по бокам (на 9-11-м сегментах). Встречаются иногда и очень слабо окрашенные, почти совсем светлые черви. Местами, особенно в Сибири, норденшельдов червь достигает 25-30 см длины; в Европейской части СССР он обычно мельче, всего 6 -12 см. Интересно, что этот вид довольно многочислен в тундрах, в районе вечной мерзлоты; недавно он был найден даже на Ляховских островах, на вечных льдах. Последний вид семейства люмбрицид, который мы упомянем, - четырехгранная эйзениелла (Eiseniella tetraedra) - один из самых мелких, длиной всего 2,5-6 см. Ее окраска серовато-коричневая, с желтоватым или оливковым оттенком; поясок с 22-23-го по 26 - 27-й сегмент; задний конец тела четырехгранный. Это земноводный вид, обитающий по берегам водоемов; иногда его можно найти и в воде у берега.
В умеренной и холодной зонах северного полушария, в частности на территории СССР, распространены почти исключительно только "настоящие" дождевые черви, т. е. представители семейства Lumbricidae, с которым мы только что познакомились. В странах южного полушария, а также в тропической и отчасти субтропической зоне северного полушария дождевые черви представлены четырьмя другими семействами, включающими в общей сложности около 1800 видов, лишь немногие из которых проникают в южные районы северного умеренного пояса. С другой стороны, некоторые виды люмбри цид, будучи завезены случайно человеком в страны южного полушария, нашли в ряде случаев подходящие для себя условия и размножились в большом количестве, успешно конкурируя с местными видами других олигохет (например, в Австралии и Новой Зеландии).
Семейство Глоссосколециды (Glossoscolecidae). Большинство из 200 с лишним видов этого семейства распространено в Южной Америке, некоторые встречаются в Центральной и даже Северной Америке, а также Африке, Юго-Восточной Азии и Австралии. В Европу проникли лишь немногие: несколько видов рода Hormogaster известны в Италии, Сицилии и Сардинии, а один вид этого семейства - Criodrilus lacuum - распространен довольно широко в странах Южной и Средней Европы, встречается и в Европейской части СССР. По внешнему виду этот довольно крупный (12-32 еж длиной, 3-5 мм толщиной), коричневато-серой окраски, иногда с зеленоватым или красноватым оттенком, червь похож на обычных дождевых червей, но по образу жизни он является водным животным и держится в иле среди корней водных растений в прибрежной части рек и озер, а также в слабосоленых водоемах, например в Азовском море. Найден также в водоемах Закавказья и в Амуре. Очень своеобразны откладываемые им коконы.
Хотя среди видов этого семейства есть и другие, ведущие водный образ жизни, большинство глоссосколецид, как и наши люмбрициды, обитает в почве. Среди них есть несколько видов, достигающих длины около 1 м, а южноамериканский Rhinodri-lus fafner даже 2 м с лишним длиной и почти 2,5 см толщиной!
Семейство Мегасколециды (Megascolecidae) - самое богатое видами семейство олигохет, распространенное в тропиках и южном полушарии. Среди его представителей есть довольно мелкие, всего 2-3 см длиной, есть и очень крупные, свыше 1 м; к этому семейству принадлежит и самый длинный земляной червь - австралийский Megascolides australis, достигающий длины 2,5 м (а по некоторым данным, даже до 3 м). Издали такого червя можно принять за крупную змею (рис. 287).
Крупные виды мегасколецид перерабатывают большое количество земли; их выбросы (рис. 288), имеющие иногда вид башенок, бывают до 20-25 см высотой. У одного из африканских видов этого семейства (Dichogaster jaculatrix) глинистые выбросы, торчащие местами в большом количестве среди опавших листьев, напоминают своим видом миниатюрные красноватые печные трубы; кстати, местные жители делают из этих выбросов трубки для курения, предварительно обжигая их.
Большинство мегасколецид по размерам и окраске слабо отличается от европейских люмбрицид; однако среди них встречаются виды довольно ярко окрашенные, иногда с кольчатым и даже мраморным рисунком. Преобладающие тона - красно-коричневые, пурпурные, сероватые; реже встречаются желтые, оливково-зеленые, синеватые, почти черные и даже перламутровые. Кроме обычного для большинства земляных червей расположения щетинок - по 4 пары на каждом сегменте, у многих видов мегасколецид наблюдается иное - в виде сплошного ряда или цепочки, опоясывающей примерно посредине каждый сегмент, причем количество щетинок в такой цепочке, т. е. на каждом сегменте, нередко бывает равно 60-70 , а в отдельных случаях доходит до 150; иногда эти ряды прерваны на спинной и брюшной сторонах.
Почти все мегасколециды - обитатели почвы; их обилие и интенсивность деятельности, как и у европейских дождевых червей, зависит от влажности почвы, температуры, обилия органических веществ и т. д.; при неблагоприятных условиях они уходят в глубокие слои и их деятельность замедляется или вовсе прекращается, например в периоды засухи. Было, однако, замечено, что один вид - Pheretima posthuma - встречается и в довольно сухих местах, причем оказывается активным на поверхности даже в засушливые периоды. Это объясняется тем, что у него выработался особый физиологический механизм для сбережения воды в теле, подобный тому, который наблюдается у насекомых: жидкие продукты обмена веществ, экскреты, не выделяются почти совсем во внешнюю среду, а попадают в кишечник, где вода из них подвергается обратному всасыванию клетками стенки кишечной трубки. Таким образом, этот вид представляет редкий среди олигохет пример высокосовершенного приспособления к наземному образу жизни.
Многие мегасколециды часто попадаются в лежащих на земле гниющих стволах деревьев, под корой, а некоторые забираются довольно высоко, до 7-8 м, на живые деревья, где держатся в углублениях коры, среди эпифитных папоротников и даже (в сырых сильно затененных джунглях) на листьях; ряд видов в связи с этим получил соответственные названия, например: древесный перионикс (Perionyx arboricola), пальмовый дихогастер (Dichogaster bolaui palmicola).
Пресноводных и литоральных (живущих в прибрежной полосе морей) видов среди представителей этого семейства мало; некоторые виды, живущие обычно во влажных почвах, встречаются иногда и в водоемах, например маленький (1,5-3 см длиной) Ocnerodrilus occidentalis, недавно найденный и у нас в Средней Азии, на рисовых полях и в некоторых водоемах. Несколько видов этого семейства было найдено в Южной Европе, а один из них, широко распространенный в Америке, Индии, Африке, Microsco lex phosphoreus, встречается и в Средней Европе. Этот червь, имеющий очень небольшие размеры (1-3,5 см), замечателен своей способностью светиться в темноте. Обычно свечение начинается, если червя потревожить, например даже при небольшом сотрясении почвы. Цвет свечения желтовато-зеленый. Светится слизь, выделяемая червем; но у погибающих червей свечение возникает и внутри тела. Большое количество этих червей наблюдалось в одной из заброшенных угольных шахт под Краковом на глубине 230 м. Зрелище было очень эффектное: в темноте двигались сотни светящихся точек. Способность светиться наблюдается и у некоторых других видов. У нас в Закавказье, в Батумском ботаническом саду, встречается много червей рода Pheretima, большинство видов которого (а их свыше 300) водится в Юго-Восточной Азии. Это очень подвижные, похожие на маленьких змеек черви, в сырую погоду быстро ползающие по поверхности земли. Найдены они и в Рионской низменности (длина их 10-15 см).
Семейство Монилигастриды (Moniligastridae). Представители его распространены в Индии, Индонезии, Индокитае, Китае и Японии, встречаются у нас на Дальнем Востоке, в южных районах Приморья. Эти, средних размеров, обычно темноокрашенные и часто отливающие радужными тонами, земляные черви попадаются иногда вместе с люмбрицидами преимущественно в сырых местах, по берегам рек и других водоемов.
Первым ученым, обратившим внимание на обилие дождевых червей в почвах, занявшимся изучением образа их жизни и пришедшим к выводу об очень большой их роли в процессах почвообразования, был Ч. Дарвин. Впервые он сообщил о своих наблюдениях над их деятельностью в небольшой статье, опубликованной еще в 1837 г., а подробно описал результаты своих многолетних исследований над ними в отдельной книге, вышедшей за год до его смерти, в 1881 г.
"Дождевые черви в истории образования земной коры играли гораздо более важную роль,- пишет Дарвин,- нежели это может казаться большинству с первого взгляда. Почти во всех влажных местностях они необыкновенно многочисленны и сравнительно с их величиной обладают весьма значительной мускульной силой. Во многих местностях Англии на площадь в акр * ежегодно выбрасывается более 10 тонн (10 516 килограммов) сухой земли, прошедшей через их тело, так что весь поверхностный слой растительной земли в течение нескольких лет проходит через их тело. Вследствие спадения стенок старых норок червей растительный слой находится хотя и в медленном, но постоянном движении, причем составные частицы перетираются друг о друга. В силу этих явлений свежие поверхностные слои постоянно подвергаются действию углекислоты, содержащейся в почве, и гумусовых кислот, роль которых в разрушении горных пород является еще более значительной. Образование гумусовых кислот обусловливается, вероятно, перевариванием полуразложившихся листьев, заглатываемых дождевыми червями. Таким образом, частицы земли, составляющие верхний растительный слой, находятся при таких условиях, которые вполне благоприятствуют их разрушению и распадению.
Черви наилучшим образом приготовляют почву для произрастания растений с мочковатыми корнями и для всякого рода семенных растений. Они периодически подвергают растительную землю действию воздуха, просеивают ее так, что в ней не остается ни одного камешка крупнее тех, которые они могут заглотить. Они равномерно перемешивают все, подобно садовнику, готовящему мелкозернистую землю для своих избранных растений. В таком виде она становится одинаково хорошо приспособленной как к сохранению влажности и поглощению растворимых веществ, так равно и для процессов нитрификации... Листья, стаскиваемые в норки червей на пищу, после того как они разорваны на мельчайшие кусочки, отчасти переваренные, отчасти смоченные жидкими выделениями кишечного канала и мочевыми выделениями, перемешиваются с большими количествами земли. Эта земля образует тогда тот темный, плодородный гумус, который почти сплошь покрывает поверхность страны хорошо обособленным слоем..."
Дарвин показал, что черви, поселившиеся под отдельными камнями, перекапывают под ними землю и, пропустив ее через свой кишечник, выбрасывают вокруг, причем прорытые ими ходы с течением времени спадаются; в результате сами камни постепенно как бы тонут, погружаясь все глубже в землю. Таким же образом происходит, по Дарвину, в течение столетий погребение остатков древних построек (почему и приходится производить раскопки для их обнаружения), хотя, конечно, в этом процессе играют свою немалую роль также вода и ветер, образующие наносы. Заключает свою замечательную книгу о дождевых червях Дарвин следующими словами: "Нельзя не удивиться, когда подумаешь о том, что весь растительный слой уже прошел через тело дождевых червей и через несколько лет снова пройдет через них. Плуг принадлежит к числу древнейших и имеющих наибольшее значение изобретений человека; но еще задолго до его изобретения почва правильно обрабатывалась червями и всегда будет обрабатываться ими. Весьма сомнительно, чтобы нашлись еще другие животные, которые в истории земной коры заняли бы столь видное место..."
Книга Дарвина дала толчок новым исследованиям, число которых продолжает увеличиваться. И если вначале многие думали, что он переоценил до некоторой степени роль червей, то теперь становится ясно, что в действительности во многих случаях значение их еще больше. Нужно только внести одну существенную поправку: в процессах почвообразования, кроме дождевых червей, участвуют многие другие беспозвоночные животные, главным образом некоторые насекомые и их личинки. И хотя в отдельных случаях, особенно там, где червей мало или вовсе нет, другие животные играют заметную роль, в общем все же первое место в процессах почвообразования среди животных принадлежит дождевым червям.
О том, что мнение Дарвина о масштабах деятельности дождевых червей не было преувеличенным, можно судить по следующим цифрам. Дарвин исходил из того, что количество червей на 1 га было равно 60-133 тыс. Позднейшие работы показали, что хотя в некоторых местах численность их может быть иногда даже несколько меньшей, но, как правило, она значительно выше, порядка 500 тыс.-2 млн. на 1 га, а в отдельных случаях достигает 5-7 млн. и даже 20 млн. Соответственно живой вес (биомасса) будет измеряться для слабо заселенных ими территорий величинами порядка от нескольких десятков килограммов до 100-120 кг на 1 га, a для тех мест, где их много,- до 2-3 т на 1 га. Если мы вспомним, что за сутки каждый червь пропускает через кишечник количество земли, равное весу его тела (а некоторые виды даже больше), то поймем, что на каждом гектаре черви перерабатывают ежесуточно от нескольких десятков килограммов земли до 2-3 т, а в среднем около четверти тонны. Расчеты, сделанные для различных мест умеренного и тропического поясов, показывают, что количество выбрасываемой червями земли таково, что оно образует за один год слой от 1 до 7 мм толщиной, в среднем около 3 мм.
Влияние деятельности дождевых червей на особенности почвы многообразно. Прежде всего, в результате перекапывания ими земли и прокладывания ходов повышается скважность почвы, она делается более рыхлой, ее объем увеличивается (на 15 - 30%). Вследствие этого облегчается доступ в глубокие слои почвы воды и атмосферного воздуха, необходимых как для корней травянистых растений и деревьев, так и для деятельности полезных микроорганизмов. На глубину около 1 м почва обычно бывает вся пронизана ходами червей, а у ряда видов их ходы идут гораздо глубже - до 4-8 м. Общая длина ходов под 1 м 2 поверхности земли нередко превышает 1 км, а в отдельных случаях достигает 8 км. По ходам червей легче проникают в глубину корни растений, особенно в более плотных почвах (рис. 289). Далее, черви постоянно перемешивают поверхностные слои, занося полуразложившиеся листья и другие органические остатки с поверхности в глубь почвы и вынося землю из нижних горизонтов на поверхность, а также откладывая экскременты внутри своих ходов. Расчеты показывают, что верхние слои почвы, по крайней мере на глубину 0,5-1 м, полностью перерабатываются червями, проходя через их кишечник, примерно за 100-200 лет.
Под влиянием червей изменяются и химические особенности почвы. В переработанной ими, смешанной с кишечной слизью и выброшенной в виде копролитов земле повышается содержание кальция, магния, аммиака, нитратов, фосфорной кислоты. Многие соединения переводятся в другую, более доступную для использования растениями форму. Благодаря выработке червями в особых известковых железках пищевода сростков кристалликов углекислого кальция - кальцита - происходит нейтрализация вредных почвенных кислот и выбрасываемая червями земля имеет щелочную реакцию. Наконец, копролиты червей, высыхая, постепенно распадаются на небольшие микропористые комочки, долго не размываемые водой. Благодаря этому переработанная червями земля приобретает стойкую мелкокомковатую структуру. Кроме того, в копролитах повышается количество полезных почвенных микроорганизмов. Все эти процессы, происходящие в почве под влиянием дождевых червей, заметно изменяют и улучшают ее состав, структуру и свойства, повышают ее плодородие.
ОБЩАЯ ХАРАКТЕРИСТИКА
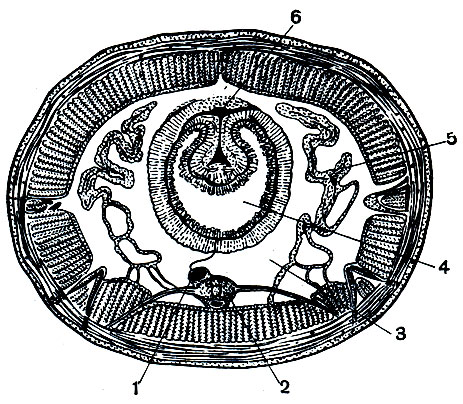
Строение и покровы . Тело малощетинковых червей (олигохет) сильно вытянутое, цилиндрическое, состоит из лежащих друг за другом колец , или сегментов . Все сегменты имеют сходное строение, т.е. для организации олигохет (и всех кольчатых червей) характерна повторяемость строения, или метамерия . Каждый сегмент, кроме самого первого, снабжен маленькими щетинками , обычно расположенными четырьмя пучками - парой боковых и парой брюшных . Передний сегмент представляет собой головную лопасть - простомиум , лишенный глаз и антенн. Он несет ротовое отверстие . Последний сегмент - анальная лопасть , или пигидиум , - несет порошицу .
Покровы олигохет представлены эпителием , образующим на поверхности тонкую эластичную кутикулу (рис. 1). Эпителий богат железистыми клетками .
Кожно-мускульный мешок . Под эпителием расположены хорошо развитые мышечные слои . Наружный слой представлен кольцевыми мышечными волокнами . За счет сокращения данного слоя тело червя утончается и вытягивается. Внутренний слой , более мощный, представлен продольными мышечными волокнами , за счет сокращения которых тело червя утолщается и укорачивается.
Рис. 1. Поперечный разрез средней части тела дождевого червя: 1 - кутикула; 2 - эпидермис; 3 - слой кольцевой мускулатуры; 4 - слой продольной мускулатуры; 5 - целомический эпителий; 6 - метанефридий; 7 - щетинка; 8 - мезентерий; 9 - брюшной сосуд; 10 - субневральный сосуд; 11 - брюшная нервная цепочка; 12 - хлорагогенные клетки; 13 - полость кишки; 14 - сосудистый плексус; 15 - тифлозоль; 16 - сосуд тифлозоля; 17 - спинной сосуд
Между кишечником и кожно-мускульным мешком находится вторичная полость тела , или целом - пространство, ограниченное собственными эпителиальными стенками мезодермального происхождения и содержащее целомическую жидкость (рис. 2). По строению целом отличается от первичной полости тела наличием целомической выстилки - собственной стенки. Выстилка образована двумя листками . Один прилегает к стенке тела, другой - к стенкам кишечника. Над кишечником и под ним оба листка срастаются, образуя брыжейку (мезентерий), которая делит целом на левую и правую стороны . Кроме этого, имеются поперечные перегородки , которые делят полость тела на камеры , соответствующие границам колец . Целом наполнен жидкостью, в которой плавают фагоциты, яйца, спермии. Целомическая жидкость, омывая внутренние органы, снабжает их кислородом и питательными веществами, а также способствует удалению продуктов обмена веществ и передвижению фагоцитов. Так же, как и жидкость, заполняющая первичную полость тела у круглых червей, целомическая жидкость может играть роль гидроскелета .
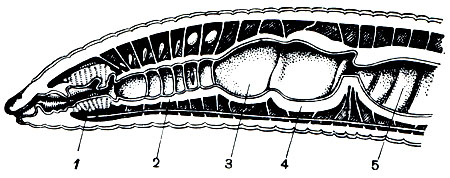
Пищеварительная система олигохет хорошо дифференцирована (рис. 3). Она начинается ротовым отверстием . Кишечник состоит из трех отделов - переднего , среднего и заднего . Наиболее дифференцирован передний отдел кишечника, состоящий из глотки, пищевода и мускульного желудка. Иногда перед желудком имеется зоб. В средней кишке для увеличения всасывательной поверхности образуется впячивание внутрь просвета кишечника - тифлозоль.
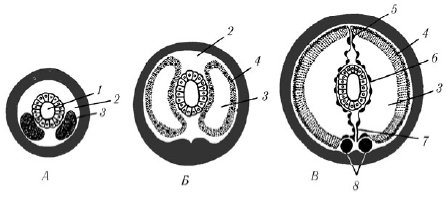
Рис. 2. Развитие целома у кольчатых червей. А-В - поперечные разрезы трех последовательных стадий развития сегмента: 1 - кишка; 2 - первичная полость тела; 3 - целом; 4 - наружная стенка целомического мешка; 5 - спинной мезентерий; 6 - внутренняя стенка целомического мешка; 7 - брюшной мезентерий; 8 - брюшные нервные стволы
Кровеносная система замкнутая и состоит из двух главных продольных сосудов - спинного и брюшного. Спинной сосуд проходит вдоль всего тела над кишкой, брюшной - под нею. Полость кровеносных сосудов представляет собой остатки первичной полости тела. Оба сосуда сообщаются кольцевыми сосудами, расположенными метамерно. Движение крови по сосудам обеспечивается пульсацией спинного сосуда и некоторых кольцевых сосудов передней части тела, называемых поэтому боковыми, или кольцевыми, сердцами. В спинном сосуде кровь течет вперед, в брюшном - назад. По кольцевым сосудам кровь движется из спинного сосуда в брюшной в передней части тела и в обратном направлении - в задних сегментах. Кровь может иметь красный цвет от железосодержащего близкого к гемоглобину позвоночных, растворенного в жидкости крови.
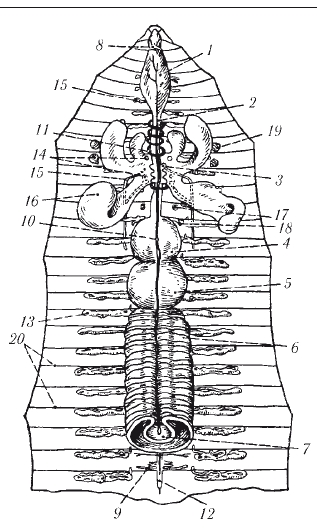
Рис. 3. Строение дождевого червя (вскрытый червь): 1 - глотка; 2 - пищевод; 3 - известковые железы; 4 - зоб; 5 - желудок; 6 - средняя кишка; 7 - спинная складка кишки (в разрезе); 8 - надглоточный нервный узел; 9 - узел брюшной нервной цепочки; 10 - спинной кровеносный сосуд; 11 - кольцевые сосуды, охватывающие глотку; 12 - брюшная нервная цепочка; 13 - метанефридии; 14 - семенники; 15 - семяпроводы; 16 - семенные мешки; 17 - яичники; 18 - яйцеводы; 19 - семяприемники; 20 - перегородки полости тела
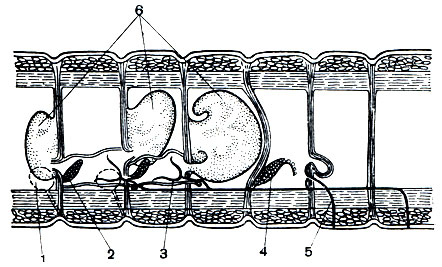
Выделительная система представлена метанефридиями. Метанефридий начинается в полости тела воронкой - нефростомом. От воронки идет проток, который проходит через перегородку, входит в соседний сегмент и открывается наружу выделительной порой в боковой стенке тела. В каждом сегменте находится пара метанефридиев - правый и левый. Воронка и проток снабжены ресничками, вызывающими движение выделяемой жидкости.
Нервная система. Центральная часть нервной системы состоит из парных мозговых ганглиев - надглоточного и подглоточного, соединенных двумя окологлоточными коннективами (нервными стволами, соединяющими разноименные ганглии). Таким образом образуется окологлоточное кольцо. В центральную часть нервной системы входит и парный брюшной нервный ствол. В каждом сегменте стволы имеют утолщения - ганглии, которые соединяются между собой перемычками - комиссурами (поперечными нервными стволами, связывающими ганглии одного сегмента). Образуется брюшная нервная цепочка, похожая на лестницу. Каждый ганглий иннервирует все органы сегмента, в котором находится.
Органы чувств у олигохет очень слабо развиты в связи с роющим образом жизни. Глаза почти всегда отсутствуют. Однако имеются светочувствительные клетки, в большом числе разбросанные в коже, что позволяет олигохетам иметь чувствительность к свету.
Половая система. Олигохеты - гермафродиты, но оплодотворение у них - перекрестное внутреннее. Половые гонады локализованы в половых сегментах. Мужские гонады - семенники - лежат в семенных капсулах, которые находятся в семенных мешках. Женская половая система представлена парой яичников, парой яйцеводов и яйцевыми мешками.
Размножение и развитие. Развитие прямое без стадии личинки. Яйца развиваются внутри яйцевого кокона, который формируется в районе пояска.
Помимо полового размножения у олигохет наблюдается бесполое размножение, похожее на бесполое размножение ресничных червей. Тело червя делится на две половины: у передней регенерирует задняя часть тела, у задней - головная часть тела.
Систематика круглых червей
Тип Круглые черви, или Первичнополостные - Nemathelminthes
Класс Нематоды - Nematoda
Подкласс Сецерненты - Secernentea
Виды Свиная аскарида - Ascaris suum
Лошадиная аскарида - Parascaris equorum
Детская острица - Enterobius vermicularis
Подкласс Аденофореи - Adenophorea
Вид Трихинелла спиральная - Trichinella spiralis
Систематика малощетинковых кольчецов
Тип Кольчатые черви - Annelida
Подтип Поисковые - Clitellata
Класс Малощетинковые - Oligochaeta
Отряд Высшие олигохеты - Lumbricomorpha
Семейство Люмбрицид - Lumbricidae
Вид Дождевой червь - Lumbricus terrestris
Часть 1
Задание 1. Рассмотрите в тубусах внешнее строение лошадиной аскариды { Parascaris equorum ). Изучите форму тела, передний, туловищный и хвостовой отделы, определите размеры (длина, толщина). Найдите отличия во внешнем строении самки от самца, определите спинную и брюшную стороны.
Исходная информация
Тело аскариды состоит из переднего, туловищного и хвостового отделов. Самки имеют удлиненное прямое веретеновидное тело. Самцы отличаются от самок меньшими размерами, их хвостовой отдел загнут крючком на брюшную сторону. На переднем конце (иногда его называют «головной») находится ротовое отверстие. Рот окружен тремя губами и занимает терминальное положение. Анальное отверстие аскарид располагается на брюшной стороне, чуть впереди от заднего конца тела. Отдел тела от анального отверстия до конца тела называется хвостом. На брюшной стороне тела самок имеется половое отверстие (рис. 1).
Рис. 1. Внешний вид лошадиной аскариды:
А - самец; Б - самка: 1 - передний конец тела; 2 - задний конец тела
Задание 2. На микропрепарате поперечного среза аскариды изучите: строение покровов, полость тела. Найдите четыре гиподермальных валика, расположенных радиально, мышечные ленты, нервные стволы, половую систему, экскреторные каналы. Сравните детали строения поперечного среза аскариды с рис. 2.
Зарисуйте поперечный срез аскариды. Обозначьте нервные тяжи в спинном и брюшном валиках гиподермы , боковые валики гиподермы с выделительными каналами, кутикулу, гиподерму, продольные мышцы, схизоцель, яйцеводы, матку, полость и стенки кишечника.
Исходная информация
Снаружи тело аскариды покрыто многослойной кутикулой, под ней заметна гиподерма. Гиподерма по бокам тела несколько утолщена, образует валики, содержащие каналы выделительной системы. Аналогичные утолщения заметны на брюшной и спинной сторонах. В них просматриваются каналы нервной системы. В первичную полость тела вдаются выросты мускульных клеток. Выросты мускульных клеток валиками гиподермы поделены на четыре ленты. Наибольшее пространство полости тела занимает кишечник. Стенка средней кишки состоит из одного слоя клеток. Матка содержит сформированные яйца аскарид. Значительное число поперечных срезов яичников и яйцеводов объясняется их большой величиной и извилистостью в полости тела .
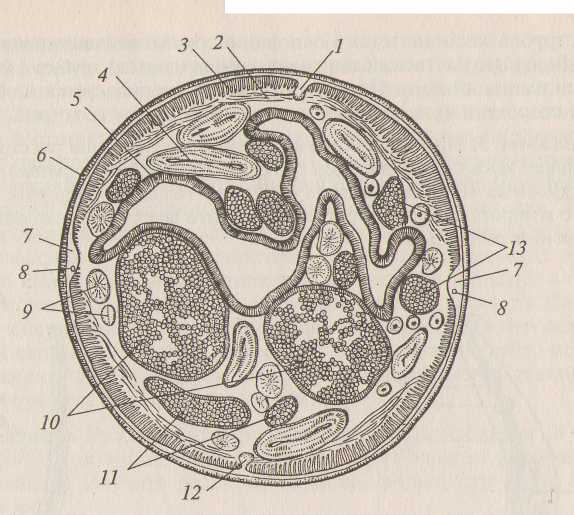
Рис. 2. Поперечный разрез самки аскариды:
1 - спинной валик гиподермы; 2 - плазматические отростки мышечных клеток; 3 - мышечные клетки; 4 - яичник в продольном разрезе; 5 - стенка кишечника; 6 - кутикула; 7 - боковой валик гиподермы; 8 - продольный канал выделительной системы; 9 - яичники, перерезанные поперек; 10 - матка; 11 - яйцеводы в продольном разрезе; 12 - брюшной валик гиподермы; 13 - яйцеводы, перерезанные поперек.
Зарисуйте внешний вид самки и самца острицы. Обозначьте рот, глотку, пищевод, бульбус, кишечник, яичник, матку, женское половое отверстие, анальное отверстие.
Исходная информация
Максимальная длина самки острицы составляет 10-12 мм, длина самцов достигает 2 - 5 мм. На переднем конце тела имеется кутикулярное обрамление - везикула. Задний участок тела заострен. Из внутренних органов просматриваются ротовое отверстие, длинный расширенный пищевод, заканчивающийся шаровидным бульбусом. За ним начинается длинный кишечник. Пищеварительная трубка заканчивается у основания хвоста анальным отверстием. Большую часть тела занимает парная извитая трубка с формирующимися яйцами. Половое отверстие располагается на брюшной стороне и чуть сдвинуто к переднему концу тела (рис. 3).
Рис. 3. Острица:
1 - рот; 2 - везикула; 3 - пищевод; 4 - бульбус; 5 - кишечник; 6 - половое отверстие; 7 - матка; 8- анальное отверстие; 9 - хвост.
Задание 4.
При большом увеличении микроскопа рассмотрите препарат мускульных волокон трихинеллы (Trichinella
spiralis
).
Среди мускульных волокон найдите овальные тельца - капсулы. Сравните микропрепараты с рис. 4. Найдите все указанные органы в строении гельминта.
Р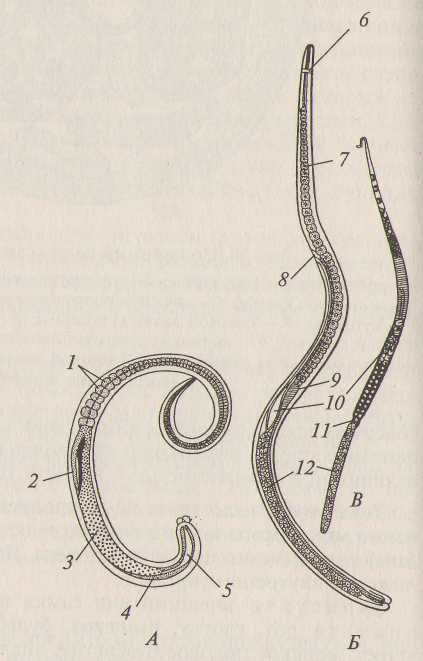
ис. 4. Внешнее и внутреннее строение трихинеллы:
А - самец; Б - самка; В - половой аппарат зрелой самки: 1,7 - клетки пищевода; 2 - семяпровод; 3 - семенник; 4 - кишечник; 5 - семенной пузырь; 6 - нервное кольцо; 8 - половое отверстие; 9 - кишка; 10 - матка с личинками; 11 - семяприемник; 12 - яичник.
Исходная информация
Рассмотрите и изучите на микропрепаратах инкапсулированных личинок трихинелл. Личинка трихинеллы располагается в маленькой капсуле и свернута в спираль. Длина маленького гельминта достигает 0,5 мм. На микропрепарате среди мускульных волокон находятся овальные тельца - капсулы. Инкапсулированные личинки сосредоточены в мышечной системе промежуточного хозяина. Таким хозяином могут быть свинья или человек. Жизнеспособными личинки остаются и через год. При проглатывании непроваренного мяса с личинками трихинелл происходит заражение. Вначале капсулы растворяются, личинки попадают в просвет кишечника и через 3 дня достигают половой зрелости. Самки после спаривания в слизистой оболочке кишечника отрождают новых личинок. Личинки проникают в кровеносное русло и разносятся по всему организму. При этом основной хозяин становится для трихинеллы промежуточным.
Задание 5. Рассмотрите строение яиц свиной аскариды. Изучите размеры, окраску, форму, структуру оболочки яиц.
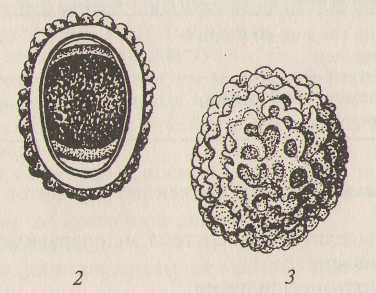
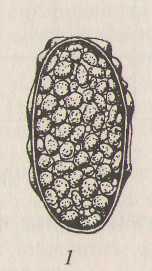
Рис 5. Яйца свиной аскариды:
1 - неоплодотворенное яйцо; 2 - оплодотворенное яйцо в оптическом разрезе; 3 - оплодотворенное яйцо с поверхности.
Задание 6. Заполните табл. 1.
Таблица 1
Черты сходства и различия в биологии круглых червей
|
Элементы сравнения |
Аскарида человеческая |
Аскарида свиная |
Острица детская |
Трихинелла |
||||||||||||||||||||||||||||||||||||||||||
|
Длина тела самок | ||||||||||||||||||||||||||||||||||||||||||||||
|
Длина тела самцов | ||||||||||||||||||||||||||||||||||||||||||||||
|
Количество яиц, продуцируемых самкой в сутки в сутки | ||||||||||||||||||||||||||||||||||||||||||||||
|
Везикула имеется | ||||||||||||||||||||||||||||||||||||||||||||||
|
Наличие или отсутствие в пищеварительной системе бульбуса | ||||||||||||||||||||||||||||||||||||||||||||||
|
Размножение яйцами или отрождение живых личинок | ||||||||||||||||||||||||||||||||||||||||||||||
|
Развитие происходит внутри одного хозяина без выхода во внешнюю среду или со сменой хозяина | ||||||||||||||||||||||||||||||||||||||||||||||
|
Развитие связано со вторичной инвазией | ||||||||||||||||||||||||||||||||||||||||||||||
|
Вызываемое заболевание хозяина |
а) протонефридиями; б) каналами с мочевым пузырем; в) метанефридиями; 2. Нерастворимые продукты обмена веществ и посторонние тела из полостной жидкости устраняются: а) протонефридиями; б) фагоцитарными клетками; в) растворяются самостоятельно; г) вообще не удаляются. 3. В спинном и брюшном валиках гиподермы у нематод располагаются истемы: а) кровеносная; б) выделительная; в) нервная; г) пищеварительная. 4. Местом откладки гликогена у нематод является: а) полостная жидкость; б) стенки каналов выделительной системы; в) стенки каналов пищеварительной системы; г) клетки мускульного слоя. 5. Органами химического чувства нематод служат: а) папиллы; б) амфиды; в) склериты; 5. Органами дыхания пресноводных олигохет - «трубочников» - являются: а) жабры наружные; б) легкие; в) поверхность тела; г) полостная жидкость. 6. Функция желудка дождевого червя: а) перетирание пищи; б) переваривание пищи; в) всасывание; г) механическое проталкивание пищи; д) накопление пищи. 7. К выделительной системе почвенных олигохет не относятся: а) метанефридии; б) хлорагогенные клетки; в) особые поры в стенке тела; г) фагоцитарные клетки. 8. У большинства олигохет половые воронки и их выводные протоки образовались из: а) мезодермы; б) эктодермы; в) эндодермы; г) выростов кишечника. Задание 6. Заполните таблицу 2. Таблица 2 Черты сходства и различия в биологии многощетинковых и малощетинковых червей
Литература: 1. Волцит О. В., Черняховский М. Е. Природа России: жизнь животных. Беспозвоночные.- М.: ООО «Фирма Издательство АСТ», 1999. – 768 с. 2. Козлов М.А., Дольник В.Р. Введение в зоологию. Простейшие. Губки. Кишечнополостные. Атлас. – СПб.: ЧеРо-на Неве, М.: Идательство МГУ, 2000. – 40 с. 3. Практикум по зоологии беспозвоночных: Учеб. Пособие для студ. Высш. Пед. Учеб. Заведений/ В.А. Шапкин, З.И. Тюмасева, И.В. Машкова, Е.В. Гуськова. – 2-е изд., испр. – М.: Издательский центр «Академия», 2005. – 208 с. 4. Шарова И.Х. Зоология беспозвоночных. – М.: Гуманит. Изд. Центр ВЛАДОС, 1999, - 592. Раздел №3 «Зоология». |