Тип аннелид, объединяющий около 12000 видов, представляет собой как бы узел родословного древа животного мира. Согласно существующим теориям, кольчатые черви ведут свое происхождение от древних ресничных червей (турбеллярная теория) или же от форм, близких к гребневикам (трохофорная теория). В свою очередь от кольчатых червей возникли в процессе прогрессивной эволюции членистоногие. Наконец, в своем происхождении аннелиды связаны общим предком с моллюсками. Все это показывает то большое значение, которое имеет рассматриваемый тип для понимания филогенеза животного мира. С медицинской точки зрения аннелиды имеют ограниченное значение. Определенный интерес представляют лишь пиявки.
Аннелиды имеют сегментированное тело. Среди самых примитивных членов метамеры почти все одинаковы. Эти метамеры получены из мезодермы, а мезодермальные органы расположены метаме - рически: мышцы, системы кровообращения, дыхательная система, осморегуляторная система и гонады. С другой стороны, органы, которые получены из энтодермы, такие как пищеварительный тракт, не являются метамерными.
Эпидермис Аннелид также образует щетинки или параподы, которые используются для локомоции. В полынке сегменты, по-видимому, похожи внешне, за исключением первых двух и последних. Первый сегмент, расположенный перед ртами, называется проломием. Рот расположен во втором сегменте, перистомие. Последний сегмент, со своей стороны, называется пигидием.
Общая характеристика типа
Тело кольчатых червей состоит из головной лопасти, сегментированного туловища и задней лопасти. Сегменты туловища на протяжении почти всего тела имеют подобные друг другу наружные придатки и сходное внутреннее строение. Таким образом, для организации аннелид характерна повторяемость строения, или метамерия.
Стена тела Аннелид состоит из нескольких слоев. Существует кутикула, секретируемая подстилающим эпидермисом. Под эпидермисом находится слой круговых мышц и слой продольных мышц. Олигочаеты имеют уменьшенные или очень маленькие щетинки и не обладают параподиями. Метамеризация целома допускает новый тип движения: перистальтическое движение. Приобретение этого транспортного средства позволило первым Аннелидам колонизировать среду, к которой другие нестерпимые организмы не имели доступа, а именно отложения, богатые органическим веществом, и могли быть источником их успеха.
По бокам тела на каждом сегменте обычно имеются наружные придатки в виде снабженных щетинками мускульных выростов - параподий - или же в виде щетинок. Эти придатки имеют значение при движении червя. Параподии в процессе филогенеза дали начало конечностям членистоногих. На головном конце тела находятся особые придатки - щупальца и палыгы.
Перистальтическое движение включает в себя гидростатический скелет и щетину. Сокращение круговых мышц сегмента вызывает его удлинение и ретракцию щетинок, которая продвигает передний конец сегмента. Затем продольные мышцы сокращаются. Это действие сопровождается расширением щетинок, которые закрепляются в субстрате, и сегмент укорачивается, вытягивая задний конец сегмента вперед. Обратите внимание, что сокращение круговых мышц приводит к растяжению продольных мышц и наоборот. Захоронение облегчается за счет приема осадка.
Волнистость тела также позволяет использовать параподы, такие как весла и плавать. Щетинки играют важную роль в локомоции и позволяют животному закрепиться, чтобы вызвать толчок. Чашевидные чаши позволяют им ползти по твердым субстратам, таким как гусеница. Пиявки также могут плавать, размахивая плоскими телами.
Развиты кожно-мускульный мешок, который состоит из кутикулы, лежащего под ней одного слоя клеток кожи и нескольких слоев мышц (см. табл. 1) и вторичная полость тела, или целом, в котором располагаются внутренние органы. Целом выстлан перитонеальным эпителием и разделен перегородками на отдельные камеры. При этом в каждом сегменте тела имеется пара целомических мешков (лишь головная и задняя лопасти лишены целома).
Дыхание в дождевых червях происходит путем диффузии через проницаемую кутикулу. Железы эпидермиса непрерывно продуцируют слизь, которая удерживает поверхность влажной и, таким образом, позволяет кислороду диффундировать внутрь животного. В дождевых червях замкнутая система кровообращения образована капиллярами, окружающими желудочно-кишечный тракт, продольными кровеносными сосудами, протекающими через тело, и боковыми сердцами которые накапливают кровь в систему кровообращения. Кровь крови в крови содержит респираторный пигмент, который повышает эффективность переноса кислорода: гемоглобин.
Целомические мешки в каждом сегменте помещаются между кишечником и стенкой тела, они заполнены водянистой жидкостью, в которой плавают амебоидные клетки.
Целом выполняет опорную функцию. Кроме того, в целомическую жидкость из кишечника поступают питательные вещества, которые затем распределяются по всему организму. В целоме накапливаются вредные продукты обмена веществ, которые удаляются выделительными органами. В стенках целома развиваются мужские и женские половые железы.
Этот пигмент находится в растворе в крови и не содержится в специализированных клетках, как у позвоночных. Морские полихеты, которые живут в норах или туннелях, испытывают больше трудностей с получением кислорода, часто имеют жабры, а поверхности параподий часто участвуют в дыхательных обменах. циркулирующей воды и помогая продлить кислород.
Дождевые черви - это детриворы, которые глотают почву и удаляют органический материал. Их мускульная глотка позволяет им проглатывать почву, которая затем измельчается в желудке. Чаша для еды перемещается по пищеварительному тракту благодаря сокращениям мышц, окружающих кишечник, которые вызывают перистальтику.
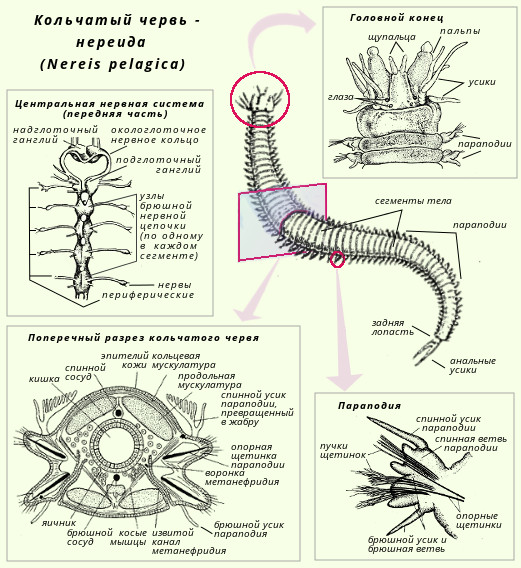
Центральная нервная система представлена надглоточным нервным узлом и брюшной нервной цепочкой. К надглоточному узлу проходят нервы от органов чувств: глаз, органов равновесия, щупалец и пальп. Брюшная нервная цепочка состоит из узлов (по одной паре в каждом сегменте тела) и стволов, связывающих узлы между собой. Каждый узел иннервирует все органы данного сегмента.
Стена пищеварительного тракта образует дорсальную складку, тифлозол, которая увеличивает поверхность контакта между проглоченной пищей и поверхностью диффузии переваренных продуктов. Поперечное сечение червячного сада, иллюстрирующего структуры, связанные с пищеварительным трактом.
Полихеты также могут быть мусорщиком, но некоторые из них являются суспендирующими, а другие - хищниками. Пиявки являются наиболее детритовыми и хищниками, некоторые из которых питаются кровью позвоночных животных, которые обычно имеют урожай и много цеек которые служат для хранения крови, из которой они сливаются, что позволяет им выжить между двумя отдаленными приемами пищи.
Пищеварительная система состоит из передней, средней и задней кишок. Передняя кишка обычно разделена на ряд отделов: глотку, пищевод, зоб и мускульный желудок. Рот находится на брюшной стороне первого сегмента тела. Задняя кишка открывается заднепроходным отверстием на задней лопасти. В стенке кишечника имеется мускулатура, обеспечивающая продвижение пищи.
Наличие целомической жидкости облегчает выделение метаболических отходов, и жидкость постоянно фильтруется метанефридами. Аннелиды имеют две метанефридии на сегмент. Каждый метанефрит открывается снаружи нефритиофором, через который выгружаются метаболические отходы, а в сточных водах отходы азота выпускаются в основном в виде мочевины. Частичная реабсорбция солей и воды вдоль трубочки, но метанефридии не являются высокоэффективной осморегуляторной системой.
Нервная система Аннелид хорошо развита с концентрацией ганглиев в передней конечности и вентральным нервным шнуром, который вступает в сегментные или периферические нервы в каждом сегменте. Сенсорные структуры уменьшаются у земляных червей, но включают в себя органы фоторецептора. Основание щетинок часто иннервируется, и эти структуры также служат в качестве механорецепторных органов. Хищные полихеты, такие как Нереис, имеют более развитые глаза.
Органы выделения - метанефридии - представляют собой парные трубчатые органы, метамерно повторяющиеся в сегментах тела. В отличие от протонефридиев они имеют сквозной выделительный каналец. Последний начинается воронкой, открывающейся в полость тела. Через воронку в нефридий поступает полостная жидкость. От воронки отходит каналец нефридия, открывающийся порой наружу. Проходя по канальцу, жидкость изменяет свой состав; в ней концентрируются конечные продукты диссимиляции, которые через наружную пору нефридия выбрасываются из организма.
Репродуктивная система хорошо развита и включает в себя несколько яичек и яичников. Во время спаривания сперма переносится с одного человека на другой и хранится в семенной емкости, где она хранится. После спаривания происходит оплодотворение яиц. Передача спермы происходит по двум бороздам, образованным складками кутикулы, что снижает шансы самооплодотворения. Самой яркой характеристикой внешней анатомии является наличие крошечной кутикулы, клителлы, с 33-го по 37-й сегмент, который, по-видимому, является созреванием гениталий.
Клителла является очень общей характеристикой олигохтетов в зависимости от родов. Переменное число сегментов, расположенных в разных положениях, имеет более или менее важное развитие. Щетинки с четырьмя парами на сегмент выходят непосредственно из стенки тела, а параподии отсутствуют.
У аннелид впервые в филогенезе животного мира появляется кровеносная система. Главные кровеносные сосуды проходят по спинной и брюшной стороне. В передних сегментах они соединены поперечными сосудами. Спинной и передние кольцевые сосуды способны ритмично сокращаться и выполняют функцию сердца. У большинства видов кровеносная система замкнутая: кровь циркулирует по системе сосудов, нигде не прерываемых полостями, лакунами или синусами. У некоторых видов кровь бесцветная, у других красная в связи с присутствием гемоглобина.
Поперечный разрез в среднем сегменте тела позволяет нам заново открыть структуру организации, которая в общих чертах напоминает о полихетской Аннелиде. Под эпидермисом, непересекающимся и покрытым кутикулой, является слой круговых мышц и, в противовес более внутренне, продольные мышцы. Маленькие протестные и ретрактивные мышцы обеспечивают мобильность щетины.
Общая полость, разделенная на уровне каждого метаметра диссекантом, заполнена жидкостью, которая может протекать наружу медианной дорзальной порой. Пищеварительный тракт, прямолинейный от рта до заднего прохода, отличается ранее в ротовой полости, а затем глоткой.
Большинство видов кольчатых червей дышит через кожу, богатую кровеносными капиллярами. У ряда морских форм имеются специализированные органы дыхания - жабры. Обычно они развиваются на параподиях или на пальпах. К жабрам подходят сосуды, несущие венозную кровь; она насыщается кислородом и в виде артериальной крови поступает в тело червя. Среди аннелид есть раздельнополые и гермафродитные виды. Половые железы расположены в полости тела.
Последний, который может быть дефлагрирован, раздувается в дорсальной части в луковице, которая играет важную роль в сборе пищи; образуя присоску, он применяется к самому большому мусору, который он вводит путем втягивания. После глотки, реснитчатый пищевод содержит калевидные железы, вмешивающиеся в ионные равновесия внутренней среды; он, наконец, диверсифицируется впоследствии в урожай и мышечную герань.
Наиболее заметной особенностью средней кишки является наличие медио-дорзальной инвагинации: тифлосолис. Этот тип желоба заполнен специализированными клетками, хлорогенными клетками, которые вмешиваются как в метаболизм углеводов, так и в экскрецию. Поэтому эти клетки играют роль накопления печени и почек.
Кольчатые черви обладают самой высокой организацией по сравнению с другими типами червей (см. табл. 1); у них впервые появляются вторичная полость тела, кровеносная система, органы дыхания, более высоко организована нервная система.
| Таблица 1. Характерные особенности различных типов червей | ||||||
| Тип | Кожно-мускульный мешок | Пищеварительная система | Кровеносная система | Половая система | Нервная система | Полость тела |
| Плоские черви | Включает слои продольных и кольцевых мышц, а также пучки спинно-брюшных и диагональных мышц | Из эктодермальной передней кишки и энтодермальной средней кишки | Не развита | Гермафродитная | Парный мозговой узел и несколько пар нервных стволов | Отсутствует, заполнена паренхимой |
| Круглые черви | Только продольные мышцы | Из эктодермальной передней и задней кйшки и энтодермальной средней кишки | То же | Раздельнополая | Окологлоточное нервное кольцо и 6 продольных стволов | Первичная |
| Из наружных кольцевых и внутренних продольных мышц | Из эктодермальной передней и задней кишки и энтодермальной средней кишки | Хорошо развита, замкнутая | Раздельнополые или гермафродиты | Парный мозговой узел, окологлоточное нервное кольцо, брюшная нервная цепочка | Вторичная | |
|
Для животных, относящихся к типу кольчатых червей, или кольчецов, характерны: Полностью закрытая система кровообращения по существу включает вентральный сосуд, дорсальный сосуд и сеть капилляров, разветвляющихся в различные органы. Кровь окрашена в красный цвет пигментом рядом с гемоглобином, эритрокроорином. Окисление происходит через стенку тела. У некоторых водных видов только жабры представляют собой дифференцированную дыхательную систему. Нервная система состоит, по существу, из брюшной нервной цепи, которая ранее была связана с церебробридиальным ганглием двумя ветвями перисофагеального ошейника. Пары сегментных нервов начинаются с уровня каждой метафоры вентрального аккорда. Органы чувств сводятся к фоторецепторным клеткам, присутствующим во всех сегментах тела и особенно обильно в головном мозге.
Кольчатые черви обитают в пресных и морских водах, а также в почве. Несколько видов обитает в воздушной среде. Главнейшими классами типа кольчатых червей являются: Кроме того, эпидермальные чувствительные корпускулы, распределенные в различных метамерах, обеспечивают тактильную и химическую чувствительность. Организация генитального аппарата, несомненно, является наибольшей оригинальностью олигохитов по отношению к полихете. Олигохтеты на самом деле всегда являются гермафродитами, а гонады представляют собой четко определенные органы с эвакуирующими каналами, независимыми от нефридий. Продукты, опосредованные сперматогонием, попадают в крупные семенные пузырьки в 9-м, 11-м и 12-м сегментах, где происходит сперматогенез. Зрелые сперматозоиды собираются в ресничных павильонах спермидуков, которые сливаются с каждой стороны, образуя пару отложенных каналов, которые открываются в 15-м сегменте.
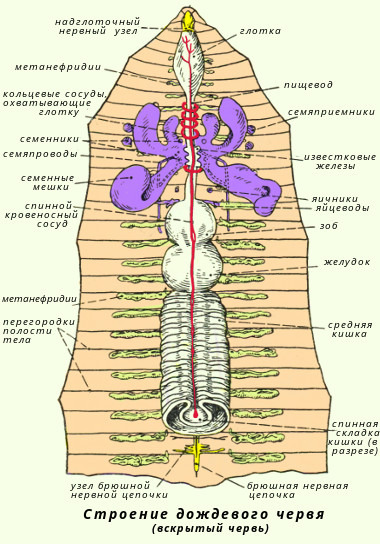
Класс многощетинковые кольчецыС точки зрения филогенеза животного мира полихеты - наиболее важная группа аннелид, так как с их прогрессивным развитием связано возникновение вышестоящих групп беспозвоночных. Тело полихет сегментировано. Имеются параподии, состоящие из спинной и брюшной ветвей, каждая из которых несет на себе усик. В мускульной стенке параподиев имеются толстые опорные щетинки, а из вершины обоих ветвей торчат пучки тонких щетинок. Функция параподий различна. Обычно это локомоторные органы, участвующие в движении червя. Иногда спинной усик разрастается и превращается в жабру. Кровеносная система у полихет хорошо развита и всегда замкнутая. Имеются виды с кожным и жаберным дыханием. Полихеты - раздельнополые черви. Обитают в морях, преимущественно в прибрежной зоне. Яичники испускают яйца, собранные флажками яйцеводов; они открываются на открытом воздухе в 14-м сегменте. Число и место различных генитальных элементов составляют важнейшие критерии системы олигохитов. Во время размножения два червяка спариваются в укусе головы, тесно связанные слизистой секретом клителлы. Сперма, выделяемая человеком, следует довольно сложному пути, прежде чем собираться сосудами, расположенными в 9-м и 10-м сегментах партнера. Яйца кладут в группы в коконе, выделяемом на уровне клителлиума. Оплодотворение происходит в коконе, поэтому оно является внешним. Характерным представителем класса может служить нереида (Nereis pelagica). Она в изобилии встречается в морях нашей страны; ведет донный образ жизни, будучи хищником, захватывает добычу челюстями. Другой представитель - пескожил (Arenicola marina) - обитает в морях, роет норы. Питается, пропуская через свой пищеварительный тракт морской ил. Дышит жабрами. Оплодотворенная яйца сегментируется по спиральному типу, и все эмбриональное развитие, которое не знает стадии трохофора, происходит в коконе. Большинство олигохтетов живут либо в грязи пресных вод, либо во влажной почве. Среди куликов некоторые виды способны к серпантистскому плаванию: это так, например, с Наакс с удлиненными щетинками. Большинство лимиколов более или менее глубоко погружаются в отложения. В лаборатории этому виду разрешалось жить в общем анаэробобии в течение 48 дней. Терриколы широко распространены во всех влажных почвах; различные виды обычно распределяются по глубине. Класс малощетинковые кольчецыМалощетинковые ведут свое происхождение от полихет. Наружными придатками тела служат щетинки, которые сидят непосредственно в стенке тела; параподий нет. Кровеносная система замкнутая; дыхание кожное. Малощетинковые кольчецы - гермафродиты. Подавляющее большинство видов - обитатели пресных вод и почвы. Характерным представителем класса может служить дождевой червь (Lumbricus terrestris). Дождевые черви обитают в почве; днем они сидят в норах, а вечером часто выползают наружу. Роясь в почве, они пропускают ее через свой кишечник и питаются содержащимися в ней растительными остатками. Дождевые черви играют большую роль в почвообразовательных процессах; они взрыхляют почву и способствуют ее аэрации; затаскивают в норы листья, обогащая почву органическими веществами; извлекают глубокие слои почвы на поверхность, а поверхностные - увлекают вглубь. Строение и размножение дождевого червя Дождевой червь имеет почти круглое в поперечном разрезе тело длиной до 30 см; насчитывают 100-180 сегментов или члеников. В передней трети тела дождевого червя находится утолщение - поясок (его клетки функционируют в период полового размножения и откладки яиц). По бокам каждого сегмента развиты по две пары коротких упругих щетинок, которые помогают животному при передвижении в почве. Тело имеет красновато-коричневую окраску, светлее на плоской брюшной и темнее на выпуклой спинной стороне. Характерной особенностью внутреннего строения является то, что у дождевых червей развиты настоящие ткани. Снаружи тело покрыто слоем эктодермы, клетки которой образуют покровную ткань. Кожный эпителий богат слизистыми железистыми клетками. Под кожей находится хорошо развитая мускулатура, состоящая из слоя кольцевых и находящегося под ним более мощного слоя продольных мышц. При сокращении кольцевых мышц тело животного вытягивается и становится тоньше, при сокращении продольных оно утолщается и раздвигает частички почвы.
Пищеварительная система начинается на переднем конце тела ротовым отверстием, из него пища поступает последовательно в глотку, пищевод (у дождевых червей в него впадают три пары известковых желез, поступающая из них в пищевод известь служит для нейтрализации кислот гниющих листьев, которыми питаются животные). Затем пища переходит в расширенный зоб, и небольшой мускульный желудок (мышцы в его стенках способствуют перетиранию пищи). От желудка почти до заднего конца тела тянется средняя кишка, в которой под действием ферментов пища переваривается и всасывается. Непереваренные остатки поступают в короткую заднюю кишку и выбрасываются наружу через заднепроходное отверстие. Дождевые черви питаются полусгнившими остатками растений, которые они глотают вместе с землей. При прохождении по кишечнику почва хорошо перемешивается с органическими веществами. Экскременты дождевых червей содержат в пять раз больше азота, в семь раз больше фосфора и в одиннадцать раз больше калия, чем обычная почва. Кровеносная система замкнутая, состоит из кровеносных сосудов. Вдоль всего тела над кишечником тянется спинной сосуд, а под ним - брюшной. В каждом сегменте их объединяет кольцевой сосуд. В передних сегментах некоторые кольцевые сосуды утолщены, их стенки сокращаются и ритмически пульсируют, благодаря этому кровь перегоняется из спинного сосуда в брюшной. Красный цвет крови обусловлен наличием в плазме гемоглобина. Для большинства кольчатых червей, в том числе и дождевых, характерно кожное дыхание, практически весь газообмен обеспечивается поверхностью тела, поэтому дождевые черви очень чувствительны к влажности почвы и не встречаются в сухих песчаных почвах, где их кожа скоро подсыхает, а после дождей, когда в почве много воды, выползают на поверхность. Выделительная система представлена метанефридиями. Метанефридий начинается в полости тела воронкой (нефростом) от которой идет проток - тоненькая петлеобразно изогнутая трубочка, открывающаяся наружу выделительной порой в боковой стенке тела. В каждом сегменте червя пара метанефридиев - правый и левый. Воронка и проток снабжены ресничками, вызывающими движение экскреторной жидкости. Нервная система имеет типичное для кольчатых червей строение (см. табл. 1), два брюшных нервных ствола, их узлы соединены между собой и образуют брюшную нервную цепочку. Органы чувств развиты очень слабо. У дождевого червя нет настоящих органов зрения, их роль выполняют отдельные светочувствительные клетки, находящиеся в кожном покрове. Там же помещаются рецепторы осязания, вкуса, обоняния. Как и гидра, дождевые черви способны к регенерации. Размножение происходит только половым путем. Дождевые черви - гермафродиты. В передней части их тела находятся семенники и яичники. Оплодотворение дождевых червей перекрестное. Во время копуляции и откладки яиц клетки пояска на 32-37-м сегменте выделяют слизь, служащую для образования яйиевого кокона, и белковую жидкость для питания развивающегося зародыша. Выделения пояска образуют своеобразную слизистую муфту. Червь выползает из нее задним концом вперед, откладывая в слизь яйца. Края муфты слипаются и образуется кокон, который остается в земляной норке. Эмбриональное развитие яиц происходит в коконе, из него выходят молодые черви. Ходы дождевых червей находятся преимущественно в поверхностном слое почвы до глубины 1 м, на зиму они спускаются до глубины 2 м. Через норки и ходы червей в почву проникают атмосферный воздух и вода, необходимые для корней растений и жизнедеятельности почвенных микроорганизмов. Через свой кишечник за сутки червь пропускает столько почвы, сколько весит его тело (в среднем 4-5 г). На каждом гектаре земли дождевые черви ежесуточно перерабатывают в среднем 0,25 т почвы, а за год они выбрасывают на поверхность в виде экскрементов от 10 до 30 т переработанной ими почвы. В Японии разводят специально выведенные породы быстро размножающихся дождевых червей и используют их экскременты для биологического метода обработки почвы. В выращиваемых на такой почве овощах и фруктах повышается содержание сахара. На важную роль дождевых червей в процессах почвообразования впервые указал Ч. Дарвин. Кольчатые черви играют значительную роль в питании донных рыб, так как местами черви составляют до 50-60 % биомассы придонных слоев водоемов. В 1939-1940 гг. из Азовского в Каспийское море был переселен червь нереис, составляющий ныне основу пищевого рациона осетровых рыб Каспийского моря.
Класс пиявкиТело сегментировано. Кроме истинной метамерии, имеется ложная кольчатость - по несколько колец в одном сегменте. Параподий и щетинок нет. Вторичная полость тела редуцировалась; вместо нее имеются синусы и щели между органами. Кровеносная система незамкнутая; кровь лишь часть своего пути проходит по сосудам и изливается из них в синусы и лакуны. Органов дыхания нет. Половая система гермафродитная. Медицинские пиявки специально разводятся и затем рассылаются в больницы. Используются, например, при лечении глазных болезней, связанных с увеличением внутриглазного давления (глаукома), при кровоизлиянии в мозг и гипертонии. При тромбозах и тромбофлебитах гирудин понижает свертываемость крови и способствует растворению тромбов. |
Кольчатые черви – билатерально-симметричные сегментированные животные.
Систематика. К типу относится 5 классов, из которых наиболее известны классы Многощетинковые (Polychaeta) – 13000 видов, Малощетинковые (Olygochaeta) – 3500 видов и Пиявки (Hirudinea) – около 400 видов.
Форма и размеры тела. Тело кольчецов в подавляющем большинстве червеобразное, в поперечном сечении круглое или овальное. Туловище имеет выраженную как внешнюю, так и внутреннюю сегментацию. В этом случае говорят об истинной метамерии. При этом метамерия распространяется и на внутреннее строение червей. У пиявок наружная сегментация не соответствует внутренней.
Размеры кольчатых червей составляют от нескольких миллиметров до 2 м (наземные формы) и даже до 3 м (морские виды).
Внешнее строение тела. У многощетинковых хорошо выражен головной отдел, несущий органы разнообразного назначения: щупальца, глазки, пальпы. У некоторых видов пальпы разрастаются в сложный ловчий аппарат. На последнем сегменте расположены одна или несколько пар чувствительных усиков. Каждый туловищный сегмент по бокам несет параподии – сложные выросты тела. Основная функция этих выростов – передвижение червя. Каждая параподия состоит из двух лопастей, внутри которых находятся многочисленные щетинки. Из них несколько более крупных, их называют ацикулы. К лопастям прикреплена пара чувствительных усиков. В состав параподии часто входит и жаберный аппарат. Параподии имеют довольно разнообразное строение.
У малощетинковых червей головной отдел выражен слабо, боковые выросты (параподии) отсутствуют. Имеются лишь сравнительно немногочисленные щетинки. На теле хорошо заметен «поясок», состоящий из утолщенных сегментов.
У пиявок на переднем и заднем конце тела расположены мощные присоски. У немногих видов по бокам имеются жаберные выросты.
Кожно-мускульный мешок. Снаружи тело кольчатых червей покрыто тонкой кутикулой, под которой лежат клетки кожного эпителия. Кожа червей богата железистыми клетками. Секрет этих клеток имеет защитное значение. У ряда видов кожные выделения идут на постройку своеобразных домиков. Щетинки червей являются производными эпителия. Под кожей залегает слой кольцевой мускулатуры, позволяющей животному изменять поперечный размер тела. Ниже располагаются продольные мышцы, служащие для изменения длины тела. У пиявок между слоями кольцевых и продольных мышц располагается слой диагональной мускулатуры. У кольчецов имеются специальные мышцы, приводящие в движение параподии, пальпы, присоски и др.
Полость тела. Пространство между стенкой тела и внутренними органами у кольчецов представляет целом – вторичную полость тела. От первичной она отличается наличием собственных эпителиальных стенок, носящих название целомического эпителия (целотелия). Целотелий покрывает продольную мускулатуру стенки тела, кишечник, мышечные тяжи и другие внутренние органы. На стенках кишки целотелий превращен в хлорагогенные клетки, выполняющие выделительную функцию. При этом целомический мешок каждого сегмента тела такой изолирован от соседних перегородками - дессепиментами. Внутри целомический мешок заполнен жидкостью, содержащей различные клеточные элементы. Целом выполняет разные функции – опорную, трофическую, выделительную, защитную и другие. У пиявок целом подвергся сильной редукции и пространство между стенкой тела и внутренними органами заполнено особой тканью - мезенхимой, в которой целом сохраняется лишь в виде узких каналов.
Средняя кишка имеет форму простой трубки, которая может усложняться. Так, у пиявок и некоторых многощетинковых кишечник имеет боковые выросты. У малощетинковых на спинной стороне кишки имеется продольная складка, которая глубоко вдается в полость кишки – тифлозоль. Эти приспособления значительно увеличивает внутреннюю поверхность средней кишки, что позволяет наиболее полно усвоить переваренные вещества. Средняя кишка имеет энтодермическое происхождение. У малощетинковых червей на границе передней и средней кишки имеется расширение – желудок. Он может быть как эктодермальным, так и энтодермальным.
Задняя кишка, которая является производным эктодермы, обычно короткая и открывается анальным отверстием.
Кровеносная система кольчатых червей замкнутая, то есть кровь везде движется по сосудам. Основные сосуды – продольные - спинной и брюшной, соединенные кольцевыми. Спинной сосуд обладает способностью к пульсации и выполняет функцию сердца. У олигохет эту функцию выполняют и кольцевые сосуды передней части тела. По спинному сосуду кровь движется сзади наперед. По кольцевым сосудам, расположенным в каждом сегменте, кровь переходит в брюшной сосуд и движется в нем спереди назад. От главных сосудов отходят более мелкие, а те в свою очередь ветвятся на мельчайшие капилляры, несущие кровь все ткани червей. У пиявок система кровеносных сосудов значительно редуцирована. Кровь движется по системе синусов – остатков целома.
Кровь большинства кольчатых червей содержит гемоглобин. Это позволяет им существовать в условиях с небольшим содержанием кислорода.
Специальных органов дыхания обычно нет, так что газообмен происходит через кожу путем диффузии. У многощетинковых червей и у некоторых пиявок имеются хорошо развитые жабры.
Выделительная система чаще всего представлена метанефридиями, которые расположены метамерно, то есть попарно в каждом сегменте. Типичный метанефридий представлен длинной извитой трубкой. Трубка эта начинается воронкой, которая открывается в целом (вторичную полость тела) сегмента, далее она пронизывает перегородку между сегментами (диссепимент) и входит в железистое метанефридиальное тело, расположенное в следующем сегменте. В этой железе трубка сильно извивается и далее открывается выделительной порой на боковой поверхности тела. Воронка и трубка покрыта ресничками, с помощью которых полостная жидкость загоняется в метанефридий. При продвижении по трубке через железу из жидкости происходит всасывание воды и различных солей, а в полости трубки остается только продукты, подлежащие удалению из организма (моча). Эти продукты и выводятся через выделительную пору. У многих видов в заднем отделе метанефридиальной трубки имеется расширение – мочевой пузырь, в котором временно скапливается моча.
У примитивных кольчатых червей органы выделения, подобно плоским червям, устроены по типу протонефридиев.
Нервная система состоит из окологлоточного кольца и брюшной нервной цепочки. Над глоткой лежит мощно развитый парный комплекс ганглиев, представляющий своеобразный головной мозг. Пара ганглиев лежит также и под глоткой. Головной мозг соединяется с подглоточными ганглиями нервными тяжами, охватывающими глотку с боков. Все это образование и называется окологлоточным кольцом. Далее, в каждом сегменте под кишечником имеется пара нервных ганглиев, которые соединены как между собой, так и с ганглиями соседних сегментов. Такая система называется брюшная нервная цепочка. От всех ганглиев отходят нервы к различным органам.
Органы чувств. На головном отделе многощетинковых червей имеются хорошо развитые органы чувств: усики и пальпы (органы осязания), глаза (иногда довольно сложные), обонятельные ямки. У некоторых форм развиты органы равновесия – статоцисты. На боковых выростах тела (параподиях) имеются усики, которые выполняют осязательную функцию.
У малощетинковых червей органы чувств развиты намного слабее, чем у многощетинковых. Имеются органы химического чувства, иногда – щупальца, статоцисты, слабо развитые глаза. В коже разбросано большое количество светочувствительных и осязательных клеток. Некоторые осязательные клетки имеют штифтик.
У пиявок в коже рассеяно множество чувствительных клеток, всегда имеются также глаза и органы химического чувства (вкусовые почки).
Половая система . Среди кольчатых червей имеются как гермафродитные, так и раздельнополые формы.
Многощетинковые черви в большинстве раздельнополы. Иногда имеет место половой диморфизм. Половые железы (гонады) образуются в целомическом эпителии. Этот процесс происходит обычно в задних сегментах червя.
У малощетинковых червей чаще встречается гермафродитизм. Половые железы расположены обычно в определенных сегментах передней части червя. Сравнительно небольшие мужские половые железы (семенники) имеют выводные протоки, которыми служат либо видоизмененные метанефридии, либо обособленные от них каналы. Более крупные женские половые железы (яичники) имеют протоки, представляющие собой измененные метанефридии. Например, при расположении яичника в 13-ом сегменте, женские половые отверстия открываются на 14-ом. Имеются также семяприемники, которые наполняются во время спаривания сперматозоидами другого червя. Пиявки в основном гермафродиты. Семенники расположены метамерно, яичников одна пара. Оплодотворение у пиявок происходит путем обмена между партнерами сперматофоров.
Размножение . Кольчатым червям присуще большое разнообразие форм размножения.
Бесполое размножение характерно для некоторых многощетинковых и малощетинковых червей. При этом происходит либо стробиляция, либо боковое почкование. Это редкий пример бесполого размножения среди высокоорганизованных животных вообще.
При половом размножении многощетинковых особи, содержащие зрелые гонады (эпитокные) переходят от ползающего или сидячего образа жизни к плавающему. А у некоторых видов половые сегменты при созревании гамет могут даже отрываться от тела червя и вести самостоятельный плавающий образ жизни. Гаметы попадают в воду через разрывы стенки тела. Оплодотворение происходит либо в воде, либо в эпитокных сегментах самки.
Размножение малощетинковых начинается с перекрестного оплодотворения. В это время два партнера прикладываются друг к другу брюшными сторонами и обмениваются спермой, которая попадает в семяприемники. После чего партнеры расходятся.
В дальнейшем на пояске выделяется обильная слизь, образующая вокруг пояска муфту. В эту муфту червь откладывает яйца. При сдвигании муфты вперед она проходит мимо отверстий семяприемников; в этот момент и происходит оплодотворение яиц. Когда муфта с оплодотворенными яйцами соскальзывает с головного конца червя, края ее смыкаются, и получается кокон, в котором происходит дальнейшее развитие. Кокон дождевых червей содержит обычно 1-3 яйца.
У пиявок размножение происходит примерно также, как и у малощетинковых червей. Коконы пиявок крупные, достигающие у некоторых видов 2 см в длину. В коконе находится у разных видов от 1 до 200 яиц.
Развитие. Зигота кольчатых червей претерпевает полное, обычно неравномерное дробление. Гаструляция происходит путем инвагинации или эпиболии.
У многощетинковых червей в дальнейшем из зародыша формируется личинка, называемая трохофорой. Она имеет реснички и довольно подвижна. Их этой личинки в последствие и развивается взрослый червь. Таким образом, у большинства многощетинковых червей развитие идет с метаморфозом. Известны виды и с прямым развитием.
Малощетинковые черви имеют прямое развитие без фазы личинки. Из яиц выходят вполне сформированные молодые черви.
У пиявок из яиц в коконе формируются своеобразные личинки, которые плавают в жидкости кокона с помощью ресничного аппарата. Таким образом, взрослая пиявка образуется путем метаморфоза.
Регенерация. Для многих кольчатых червей характерна развитая способность регенерировать утраченные части тела. У некоторых видов целый организм может восстановиться всего из нескольких сегментов. Однако у пиявок регенерация выражена очень слабо.
Питание. Среди многощетинковых червей встречаются как хищники, так и растительноядные виды. Известны также факты каннибализма. Часть видов питаются органическими остатками (детритофаги). Малощетинковые черви являются в основном детритофагами, но встречаются также и хищники.
Малощетинковые черви большей частью являются почвенными обитателями. В богатых перегноем почвах численность, например червей энхитреид достигает 100-200 тысяч на квадратный метр. Обитают малощетинковые они так же в пресных, солоноватых и соленых водоемах. Водные жители населяют в основном поверхностные слои грунта и растительность. Часть видов – космополиты, есть и эндемики.
Пиявки населяют пресные водоемы. Немногие виды обитают в морях. Некоторые перешли к наземному образу жизни. Эти черви ведут либо засадный образ жизни, либо активно разыскивают своих хозяев. Однократное кровососание обеспечивает пиявок пищей на многие месяцы. Среди пиявок нет космополитов; они приурочены к определенным географическим зонам.
Палеонтологические находки кольчатых червей очень немногочисленны. Большее разнообразие в этом отношении представляют многощетинковые. От них сохранились не только отпечатки, но и во многих случаях остатки трубок. На этом основании предполагается, что все основные группы этого класса были представлены уже в палеозое. Достоверных остатков малощетинковых червей и пиявок к настоящему времени не обнаружено.
Происхождение. В настоящее время наиболее правдоподобна гипотеза происхождения кольчатых червей от паренхиматозных предков (ресничных червей). Наиболее примитивной группой принято считать многощетинковых. Именно от этой группы вероятнее всего ведут свое происхождение малощетинковые, а от последних выделилась группа пиявок.
Значение. В природе кольчатые черви имеют огромное значение. Населяя различные биотопы, эти черви включаются в многочисленные пищевые цепочки, служа пищей огромному количеству животных. Наземные черви играют ведущую роль в почвообразовании. Перерабатывая растительные остатки, они обогащают почву минеральными и органическими веществами. Их ходы способствуют улучшению газообмена почвы и ее дренажу.
В практическом плане ряд видов дождевых червей используется в качестве производителей биогумуса. Червь – энхитрей используется в качестве корма для аквариумных рыбок. Энхитреев размножают в огромных количествах. В тех же целях добывают в природе червя трубочника. Медицинских пиявок в настоящее время используют для терапии некоторых заболеваний. В некоторых тропических странах в пищу употребляют палоло – половые (эпитокные) сегменты червей, отделившихся от передней части животных и всплывших к поверхности воды.
Общая характеристика типа Членистоногие .
Членистоногие – двусторонне-симметричные сегментированные животные с метамерно расположенными членистыми конечностями. Это самая богатая видами и разнообразная группа животных.
Систематика. Тип членистоногие разделяют на несколько подтипов.
Подтип Жабернодышащие (класс Ракообразные)
Подтип Трилобиты (вымершая группа)
Подтип Хелицеровые (класс Меростомовые, класс Паукообразные)
Подтип Первичнотрахейные
Подтип Трахейнодышащие (класс Многоножки, класс Насекомые).
К классу Меростомовые относятся современные мечехвосты и вымершие ракоскорпионы . К подтипу Первичнотрахейные относятся мелкие (до 8 см) тропические животные, которые по строению занимают промежуточное положение между кольчатыми червями и членистоногими. Эти группы животных здесь рассматриваться не будут.
Размеры тела. Длина тела членистоногих колеблется от 0,1 мм (некоторые клещи) до 90 см (мечехвосты). Наземные членистоногие достигают 15-30 см. Размах крыльев некоторых бабочек превышает 25 см. Вымершие ракоскорпионы достигали 1,5 м длины, а размах крыльев ископаемых стрекоз доходил до 90 см.
Внешнее строение . Тело большинства членистоногих состоит из головы, груди и брюшка. В состав перечисленных отделов входит разное число сегментов.
Голова , сегменты которой соединены неподвижно, несет на себе ротовые органы и органы чувств. Голова подвижно ил неподвижно соединена со следующим отделом – грудью.
Грудной отдел несет на себе ходильные конечности. В зависимости от числа грудных сегментов конечностей может быть разное количество. У насекомых к груди крепятся также и крылья. Сегменты груди соединяются между собой подвижно или неподвижно.
Брюшко вмещает в себя большую часть внутренних органов и состоит чаще всего их нескольких сегментов, подвижно соединенных между собой. На брюшке могут располагаться конечности, и другие придатки.
Ротовой аппарат у членистоногих очень сложно устроен. В зависимости от способа питания он может иметь самое разнообразное строение. Части ротового аппарата по большей части представляют собой сильно видоизмененные конечности, приспособленные для употребления практически любой пищи. В состав аппарата могут входить 3-6 пар конечностей.
Покровы. Кутикула, состоящая из хитина, является производным погруженного эпителия – гиподермы. Хитин выполняет опорную и защитную функцию. Кутикула может пропитываться карбонатом кальция, становясь при этом очень прочным панцирем, как это происходит, например, у ракообразных. Таким образом, у членистоногих покровы тела представляют собой наружный скелет. Подвижное соединение твердых участков кутикулы обеспечивается наличием перепончатых участков. Кутикула членистоногих не эластична и не может растягиваться в процессе роста животных, поэтому они периодически сбрасывают старую кутикулу (линяют) и, пока новая кутикула не затвердела, увеличиваются в размерах.
Полость тела. В процессе эмбрионального развития у членистоногих закладываются целомические мешки, но в дальнейшем они разрываются и полость их сливаются с первичной полостью тела. Так образуется смешанная полость тела – миксоцель.
Мускулатура представлена отдельными мышечными пучками, которые не образуют сплошного мышечного мешка. Мышцы крепятся как непосредственно к внутренней стенке сегментов тела, так и к внутренним их отросткам, составляющим внутренний скелет. Мускулатура у членистоногих поперечнополосатая .
Пищеварительная система у членистоногих в общем случае состоит из переднего, среднего и заднего отдела кишечника. Передний и задний отдел выстлан изнутри тонкой хитиновой кутикулой. В зависимости от типа питания строение кишечника крайне разнообразно. В ротовую полость открываются слюнные железы, которые очень часто вырабатывают ряд ферментов, в том числе и пищеварительных. Анальное отверстие открывается, как правило, на заднем конце тела.
Выделительная система у первичноводных членистоногих (ракообразных) представлена специальными железами, расположенными в головной части тела. Протоки этих желез открываются у основания антенн (усиков). У наземных членистоногих выделительная система представлена так называемыми мальпигиевыми сосудами – трубками, которые с одного конца слепо замкнуты, а другим концом открываются в кишечник на границе среднего и заднего отделов. Эти трубочки, размещаются в полости тела, и, омываясь гемолимфой, всасывают из нее продукты распада и выводят их в кишку.
Дыхательная система устроена достаточно разнообразно. У ракообразных имеются настоящие жабры . Они представляют собой ветвистые выросты на конечностях, покрытые тонкой хитиновой кутикулой, через которые и происходит газообмен. Некоторые ракообразные приспособились жить на суше (например, мокрицы).
У пауков и скорпионов органы дыхания представлены листовидными легкими , которые открываются наружу отверстиями (стигмами). Внутри легочный мешок имеет многочисленные складки. Кроме легочного мешка у некоторых пауков имеется система трахейных трубок, которые практически не ветвятся.
У клещей, многоножек и насекомых дыхательная система представлена трахеями , которые открываются наружу отверстиями (дыхальца, стигмы). Трахеи сильно ветвятся и проникают во все органы и ткани. Трахея имеет тонкую хитиновую выстилку и укреплена изнутри хитиновой спиралью, которая не позволяет трубочке спадаться. Кроме того, у летающих насекомых имеются расширения – воздушные мешки, наполняющиеся воздухом и уменьшающие удельный вес животного. Вентиляция в трахейной системе идет как пассивно (диффузия), так и активно (изменение объема брюшка).
У некоторых личинок насекомых имеются особые органы дыхания – трахеальные жабры . Газообмен у таких членистоногих идет путем диффузии.
У некоторых клещей дыхательная система отсутствует, и газообмен идет через всю поверхность тела.
Кровеносная система у всех членистоногих незамкнута я, то есть не везде кровь течет по сосудам. Под хитиновым покровом спины имеется сердце, от которого отходят кровеносные сосуды. Однако на некотором расстоянии от сердца стенки сосудов исчезают, и дальнейший свой путь кровь совершает по щелям между внутренними органами. Затем она попадает в сердце через отверстия, которые называются остии. У ракообразных и клещей сердце мешковидное, а у скорпионов, пауков и насекомых оно многокамерное. У некоторых клещей кровеносная система может отсутствовать.
Кровь у подавляющего большинства членистоногих бесцветна и обычно называется гемолимфой. Это достаточно сложная жидкость: она состоит как из собственно крови, так и полостной жидкости. В связи с отсутствием специальных пигментов, гемолимфа практически не может активно участвовать в процессе газообмена. Гемолимфа некоторых насекомых (листоедов, божьих коровок) содержит довольно токсичные вещества, и может играть защитную роль.
Жировое тело. У наземных членистоногих имеется запасающий орган – жировое тело, расположенное между внутренностями. Жировое тело принимает участие в регуляции водного обмена.
Нервная система. В общем случае у членистоногих нервная система построена по типу кольчатых червей. В ее состав входят парный надглоточный ганглий, окологлоточное нервное кольцо и брюшная нервная цепочка. От ганглиев цепочки отходят периферические нервы. Особого развития достигает надглоточный ганглий у насекомых, у которых обычно говорится о наличии головного мозга. Часто наблюдается концентрация ганглиев брюшной нервной цепочки и образование за счет их слияния крупных нервных узлов. Такая концентрация часто связана с уменьшением числа сегментов (слиянием их между собой). Например, у клещей, утративших сегментацию, брюшная цепочка превращается в общую нервную массу. А у многоножек, тело которых состоит из множества одинаковых сегментов, нервная цепочка очень типична.
Органы чувств у большинства членистоногих достигают высокого развития.
Органы зрения расположены на голове и часто представлены сложными (фасеточными глазами), которые занимают у некоторых насекомых большую часть поверхности головы. У многих ракообразных фасеточные глаза сидят на стебельках. Кроме того, у насекомых и паукообразных имеются простые глазки. Непарный лобный глазок характерен для некоторых ракообразных.
Органы осязания представлены различными щетинками и волосками, расположенных на теле и конечностях.
Органы обоняния и вкуса. Большая часть обонятельных окончаний расположено на усиках и челюстных щупиках насекомых, а так же на антеннуллах ракообразных. Обоняние у насекомых развито очень хорошо: достаточно 100 молекул феромона на 1 см 2 воздуха, выделенного самкой шелкопряда, чтобы самец начал поиск партнерши. Органы вкуса у насекомых располагаются как на ротовых конечностях, так и на концевых члениках ног.
Органы равновесия . У ракообразных в основном членике антеннул расположен статоцист – впячивание кутикулы, усаженное изнутри чувствительными волосками. В этой полости обычно находятся мелкие песчинки, играющие роль статолитов.
Органы слуха. У некоторых насекомых хорошо развиты так называемые тимпанальные органы, воспринимающие звуки. Например, у кузнечиков они располагаются на основаниях голеней передних ног. Как правило, те насекомые, которые способны воспринимать звуки, способны также их издавать. К таким относятся многие прямокрылые, некоторые жуки, бабочки и др. Для этого у насекомых есть специальные приспособления, расположенные на теле, крыльях и конечностях.
Прядильные железы. Для некоторых членистоногих характерно наличие прядильных желез. У пауков они располагаются в брюшке и открываются паутинными бородавками на кончике брюшка. Пауки используют свою паутину чаще всего для охоты и постройки убежищ. Эта нить - одна из самых прочных в природе.
У личинок ряда насекомых прядильные железы находятся в переднем отделе тела и открываются около ротового отверстия. Их паутинка идет большей частью на постройку убежища или кокона.
Половая система. Членистоногие являются раздельнополыми животными, которым очень часто присущ половой диморфизм. Самцы отличаются от самок более яркой окраской и зачастую меньшей величиной. У самцов насекомых усики намного сильнее развиты.
Половая система самок состоит из желез – яичников, яйцеводов и влагалища. Сюда же относятся придаточные железы и семяприемники. Из наружных органов может присутствовать яйцеклад различного строения.
У самцов органы размножения представлены семенниками, выносящими протоками и придаточными железами. У ряда форм имеются различно устроенные копулятивные органы.
Полиморфизм. В колониях общественных насекомых имеются особи, отличающиеся друг от друга по строению, по физиологии и поведению. В гнездах пчел, муравьев и термитов присутствует, как правило, только одна самка, способная откладывать яйца (матка или царица). Самцы в колонии присутствуют либо постоянно, либо появляются по мере истощения у матки запаса сперматозоидов от предыдущего спаривания. Все остальные особи называются рабочими, которые являются самками с угнетенной половой функцией. У термитов и муравьев рабочие особи разделены на касты, каждая из которых выполняет определенную функцию (сбор пищи, защита гнезда и т.д.). Появление в гнезде самцов и полноценных самок происходит только в определенное время.
Биология размножения. Как уже говорилось, членистоногие являются раздельнополыми животными. Однако среди них нередки случаи партеногенеза (тли, дафнии). Иногда спариванию предшествует ритуал ухаживания, и даже драки самцов за самку (у жуков-оленей). После спаривания самка иногда съедает самца (богомолы, некоторые пауки).
Чаще всего происходит откладка яиц группами или по одному. У некоторых членистоногих развитие яиц и личинок происходит в организме самки. В этих случаях имеет место живорождение (скорпионы, некоторые мухи). В жизни многих видов членистоногих имеет место забота о потомстве.
Плодовитость членистоногих колеблется в очень широких пределах и зависит очень часто от экологических условий. У некоторых тлей, например, самки откладывают только одно зимующее яйцо. Матка медоносной пчелы способна отложить до 3000 яиц в день, а матка термитов до 30000 в день. За свою жизнь эти насекомые откладывают миллионы яиц. В среднем же плодовитость составляет несколько десятков или сотен яиц.
Развитие . У большинства членистоногих развитие происходит с метаморфозом, то есть с превращением. Из яйца выходит личинка, которая после нескольких линек личинка превращается во взрослое животное (имаго). Часто личинка сильно отличается от имаго как по строению, так и по образу жизни.
В цикле развития ряда насекомых присутствует фаза куколки (бабочки, жуки, мухи). В этом случае говорят о полном метаморфозе . У других (тли, стрекозы, клопы) такой фаза нет, и метаморфоз этих насекомых называется неполным .
У некоторых членистоногих (пауков, скорпионов) развитие прямое. В этом случае из яиц выходят вполне сформированные молодые животные.
Продолжительность жизни членистоногих исчисляется обычно несколькими неделями или месяцами. В ряде случаев развитие затягивается на годы. Например, личинки майских жуков развиваются около 3 лет, жуков оленей – до 6 лет. У цикад личинки живут в почве до 16 лет и только после этого превращаются во взрослых цикад. Личинки поденок живут в водоемах 1-3 года, а взрослое насекомое живет всего несколько часов, за которые успевает спариться и отложить яйца.
Распространение и экология . Представители типа членистоногие встречаются практически в любом биотопе. Они встречаются на суше, в пресных и соленых водоемах, а так же в воздухе. Среди членистоногих есть как широко распространенные виды, так и эндемики. К первым можно отнести бабочку капустную белянку, рачков – дафний, почвенных клещей. К эндемичным видам относятся, например, крупная и очень красивая бабочка брамея , которая встречается только в Колхидской низменности.
Распространение отдельных видов ограничивают различные экологические факторы.
Из абиотических факторов самыми важными являются температура и влажность. Температурные пределы активного существования членистоногих лежат в пределах от 6 до 42°С. При понижении или повышении температуры животные впадают в состояние оцепенения. Разные фазы развития членистоногих по-разному переносят температурные колебания.
Влажность среды так же во многом определяет возможность существования членистоногих. Чрезмерно низкая влажность среды, как и высокая, может привести к гибели. Для водных членистоногих наличие жидкой влаги является необходимым условием для активного существования.
На распространение членистоногих большое влияние оказывает и человеческая деятельность (антропогенное влияние ). Изменение экологических условий ведет к смене видового состава. В результате промышленной и сельскохозяйственной деятельности человека одни виды исчезают, а другие виды чрезвычайно бурно размножаются, становясь при этом вредителями.
Происхождение. Большинство исследователей сходятся в том, что членистоногие произошли от предков, близких к кольчатым червям. Предполагается, что ракообразные, хелицеровые и вымершие трилобиты произошли от кольчецов одним общим корнем, а многоножки и насекомые - другим.
Палеонтологический материал по членистоногим очень обширен. Благодаря хитиновой кутикуле остатки их достаточно хорошо сохраняются в окаменевшем виде. Исключительно хорошо сохранились наземные членистоногие и в янтаре. Однако, не смотря на это точно проследить эволюцию членистоногих сложно: отдаленные предки членистоногих в геологических слоях не сохранились. Поэтому основными методами исследования этого вопроса являются сравнительно-анатомические и сравнительно-эмбриологические.
В практической деятельности человека принято различать полезные и вредные виды.