Общая характеристика. Тип кольчатые черви, или кольчецы, охватывает около 9000 видов высших червей. Эта группа животных имеет большое значение для понимания путей филогенеза высших беспозвоночных. Кольчатые черви обладают более высокой организацией по сравнению с плоскими и круглыми червями. Они обитают в морских и пресных водах, а также в почве. Тип делится на несколько классов. Ознакомимся с представителем класса малощетинковых (дождевой червь).
Тело кольчецов состоит из сегментов. Сегменты туловища внешне одинаковы. Каждый сегмент, кроме переднего, несущего ротовое отверстие, снабжен маленькими щетинками. Это последние остатки исчезнувших пара подий.
У кольчатых, червей имеется хорошо развитый кожно-мускульный мешок, состоящий из одного слоя эпителия и двух слоев мышц: наружного слоя кольцевых мышц и внутреннего слоя, образованного продольными мышечными волокнами.
Между кожно-мускульным мешком и кишками находится вторичная полость тела, или целом, которая образуется во время эмбриогенеза внутри разрастающихся мезодермальных мешков. Морфологически она отличается от первичной полости наличием эпителиальной выстилки, прилегающей с одной стороны к стенке тела, а с другой - к стенкам пищеварительной трубки. Листки выстилки срастаются над и под кишками, а образующейся из них брыжейкой делят целом на правую и левую стороны. Поперечные перегородки делят полости тела на камеры, соответствующие границам наружных колец. Целом наполнен жидкостью.
Появление вторичной полости тела обеспечивает кольчатым червям более высокий, чем у других червей, уровень процессов жизнедеятельности. Целомическая жидкость, омывая органы тела, наряду с кровеносной системой снабжает их кислородом, а также способствует удалению продуктов жизнедеятельности и передвижению фагоцитов.
Пищеварительная система начинается ротовым отверстием на переднем конце тела и заканчивается сзади анальным. Кишка состоит из трех отделов: переднего (эктодермального), среднего (энтодермального) и заднего (эктодермального). Передняя кишка часто бывает представлена несколькими отделами; ротовой полостью и мускулистой глоткой. В стенке глотки помещаются так называемые слюнные железы. У некоторых хищных кольчатых червей имеются кутикулярные «зубы», служащие для схватывания добычи. В стенке кишки появляется слой мышц, что обеспечивает ее самостоятельную перистальтику. Средняя кишка переходит в короткую заднюю, заканчивающуюся анальным отверстием.
Дыхательная система. У дождевого червя газообмен происходит через богатую кровеносными сосудами кожу, у некоторых морских кольчецов имеются жабры.
Выделительная система. В каждом членике дождевого червя имеется парный орган выделительной системы, состоящий из воронки и извитого канальца. Продукты жизнедеятельности из полости тела попадают в воронку. От воронки идет каналец, который входит в соседний сегмент, образует несколько петель и открывается наружу выделительной порой в боковой стенке тела. Как воронка, так и каналец снабжены ресничками, вызывающими движение выделяемой жидкости. Такие органы выделения называются метанефридиями.
Кровеносная система. У большинства кольчатых червей она замкнутая, состоит из брюшного и спинного сосудов, на переднем и заднем концах тела переходящих друг в друга. В каждом сегменте кольцевой сосуд соединяет спинной и брюшной сосуды. Кровь движется по сосудам благодаря ритмичным сокращениям спинного и передних кольцевых сосудов.
Нервная система значительно усложнена по сравнению с плоскими и круглыми червями. Вокруг глотки имеется окологлоточное нервное кольцо, состоящее из надглоточного и подглоточного узлов, соединенных перемычками. На брюшной стороне находятся два нервных ствола, имеющих в каждом сегменте утолщения - ганглии, которые соединяются между собой перемычками. У многих видов кольчецов происходит сближение правого и левого нервных стволов, вследствие чего образуется брюшная нервная цепочка.
Из органов чувств у кольчатых червей есть усики, глазки, органы равновесия, которые чаще расположены на головной лопасти.
Регенерация. Дождевой червь так же, как гидра и ресничные черви, способен к регенерации, т. е. восстановлению утраченных частей тела. Если дождевого червя разрезать на две части, то у каждой из них восстановятся недостающие органы.
Половая система состоит из женских гонад (яичников), которые представляют собой комплекс половых клеток, окруженных эпителием, и мужских гонад (семенников), лежащих внутри объемистых семенных мешков.
Дождевые черви - гермафродиты, но среди кольчецов встречаются и раздельнополые формы. На теле дождевого червя имеется поясок, вырабатывающий слизь, из которой формируется кокон. В него откладываются яйца и там же происходит их развитие.
Развитие. У дождевого червя развитие прямое, но у некоторых кольчецов из оплодотворенного яйца развивается личинка, т. е. развитие происходит с превращением.
Таким образом, кольчатые черви обладают рядом прогрессивных признаков, к числу которых относится появление сегментации, целома, кровеносной и дыхательной систем, а также повышение организации выделительной и нервной систем.
Значение кольчатых червей в природе. Многие из многощетинковых червей служат основной пищей рыб, в связи с чем большое значение имеют в круговороте веществ в природе. Например, один из видов кольчатых червей - нереис, живущий в Азовском море, служит пищей промысловых рыб. Советскими зоологами он был акклиматизирован в Каспийском море, здесь интенсивно размножался и ныне является важной составной частью в питании осетровых рыб. Многощетинковый червь, называемый туземцами Полинезии «палоло», употребляется ими в пищу.
Дождевые черви питаются растительными остатками, находящимися в почве, которую пропускают через кишки, оставляя на поверхности кучки экскрементов, состоящих из земли. Этим они способствуют перемешиванию и, следовательно, разрыхлению почвы, а также обогащению ее органическими веществами, улучшению водного и газового баланса почвы. Еще Ч. Дарвин отмечал полезное влияние кольчатых червей на плодородие почвы.
Кольчатые черви известны со среднего кембрийского периода. Считается, что они произошли от низших плоских червей, так как определенные особенности их строения свидетельствуют об сходстве этих групп животных. Многощетинковых червей выделяют как основной класс типа кольчатых. От них позднее в ходе эволюции в связи с переходом к наземному и пресноводному образу жизни произошли малощетинковые, которые дали начало пиявкам.
Все кольчатые черви имеют характерное строение. Основная характеристика: их двусторонне-симметричное тело можно условно разделить на головную лопасть, сегментированное туловище и заднюю (анальную) лопасть. Число сегментов туловища может достигать от десятков до нескольких сотен. Размеры варьируют от 0,25 мм до 5 м. На головном конце кольчецов размещены органы чувств: глаза, обонятельные клетки и ресничные ямки, реагирующие на действие различных химических раздражителей и воспринимающие запахи, а также органы слуха, имеющие строение подобно локаторам. Органы чувств могут быть расположены и на щупальцах. Тело кольчатых червей разделено на сегменты в виде колец. Каждый сегмент в определенном смысле представляет собой самостоятельную часть целого организма, так как целом (вторичная полость тела) разделен перегородками на сегменты в соответствии с наружными кольцами. Поэтому этому типу присвоено такое название – «кольчатые черви». Значение такого деления тела огромно. При повреждении червь утрачивает содержимое нескольких сегментов, остальные остаются сохранными, и животное быстро регенерирует. Метамерия (сегментация) внутренних органов, и соответственно, систем органов кольчатых червей обусловлена сегментацией их тел. Внутренней средой организма кольчецов является целомическая жидкость, которая заполняет целом в кожно-мускульном мешке, состоящем из кутикулы, кожного эпителия и двух групп мышц – кольцевых и продольных. В полости тела поддерживается биохимическое постоянство внутренней среды, и могут реализоваться транспортная, половая, выделительная, опорно-двигательная функции организма. У более древних многощетинковых червей на каждом сегменте тела имеются параподии (парные примитивные конечности со щетинками). Одни виды червей передвигаются за счет сокращения мышц, а другие, используя параподии.
Ротовое отверстие находится на брюшной стороне первого сегмента. Пищеварительная система кольчатых червей сквозная. Кишечник делится на переднюю, среднюю и заднюю кишку. Кровеносная система кольчатых червей замкнутая, состоит из двух основных сосудов – спинного и брюшного, которые соединены между собой кольцевыми сосудами наподобие артерий и вен. Кровь червей этого типа может быть разного цвета у различных видов: красная, зеленая или прозрачная. Это зависит от химической структуры дыхательного пигмента в крови. Процесс дыхания осуществляется всей поверхностью тела червя, но у некоторых видов червей уже имеются жабры. Выделительная система представлена парными протонефридиями, метанефридиями или миксонефридиями (прообразы почек), имеющимися в каждом сегменте. Нервная система кольчатых червей включает крупный нервный ганглий (прообраз головного мозга) и брюшную нервную цепочку из более мелких ганглиев в каждом сегменте. Большинство кольчатых червей раздельнополы, но у некоторых вторично развился гермафродитизм (как у дождевого червя и пиявки). Оплодотворение осуществляется внутри организма либо во внешней среде.
сквозная. Кишечник делится на переднюю, среднюю и заднюю кишку. Кровеносная система кольчатых червей замкнутая, состоит из двух основных сосудов – спинного и брюшного, которые соединены между собой кольцевыми сосудами наподобие артерий и вен. Кровь червей этого типа может быть разного цвета у различных видов: красная, зеленая или прозрачная. Это зависит от химической структуры дыхательного пигмента в крови. Процесс дыхания осуществляется всей поверхностью тела червя, но у некоторых видов червей уже имеются жабры. Выделительная система представлена парными протонефридиями, метанефридиями или миксонефридиями (прообразы почек), имеющимися в каждом сегменте. Нервная система кольчатых червей включает крупный нервный ганглий (прообраз головного мозга) и брюшную нервную цепочку из более мелких ганглиев в каждом сегменте. Большинство кольчатых червей раздельнополы, но у некоторых вторично развился гермафродитизм (как у дождевого червя и пиявки). Оплодотворение осуществляется внутри организма либо во внешней среде.
Значение кольчатых червей очень велико. Следует отметить их важную роль в пищевых цепях в естественной среде обитания. В хозяйстве люди стали использовать морские виды кольчецов как кормовую базу для выращивания ценных промысловых видов рыб, к примеру осетровых. Дождевой червь издавна используется в качестве наживки при рыбной ловле, как корм для птиц. Польза от дождевых червей огромна, так как они аэрируют и разрыхляют почву, что увеличивает урожаи сельскохозяйственных растений. В медицине широко применяются пиявки при гипертонии, повышенной свертываемости крови, так как они выделяют особое вещество (гирудин), обладающее свойством уменьшать свертываемость крови и расширять кровеносные сосуды.
Тип аннелид, объединяющий около 12000 видов, представляет собой как бы узел родословного древа животного мира. Согласно существующим теориям, кольчатые черви ведут свое происхождение от древних ресничных червей (турбеллярная теория) или же от форм, близких к гребневикам (трохофорная теория). В свою очередь от кольчатых червей возникли в процессе прогрессивной эволюции членистоногие. Наконец, в своем происхождении аннелиды связаны общим предком с моллюсками. Все это показывает то большое значение, которое имеет рассматриваемый тип для понимания филогенеза животного мира. С медицинской точки зрения аннелиды имеют ограниченное значение. Определенный интерес представляют лишь пиявки.
Общая характеристика типа
Тело кольчатых червей состоит из головной лопасти, сегментированного туловища и задней лопасти. Сегменты туловища на протяжении почти всего тела имеют подобные друг другу наружные придатки и сходное внутреннее строение. Таким образом, для организации аннелид характерна повторяемость строения, или метамерия.
По бокам тела на каждом сегменте обычно имеются наружные придатки в виде снабженных щетинками мускульных выростов - параподий - или же в виде щетинок. Эти придатки имеют значение при движении червя. Параподии в процессе филогенеза дали начало конечностям членистоногих. На головном конце тела находятся особые придатки - щупальца и палыгы.
Развиты кожно-мускульный мешок, который состоит из кутикулы, лежащего под ней одного слоя клеток кожи и нескольких слоев мышц (см. табл. 1) и вторичная полость тела, или целом, в котором располагаются внутренние органы. Целом выстлан перитонеальным эпителием и разделен перегородками на отдельные камеры. При этом в каждом сегменте тела имеется пара целомических мешков (лишь головная и задняя лопасти лишены целома).
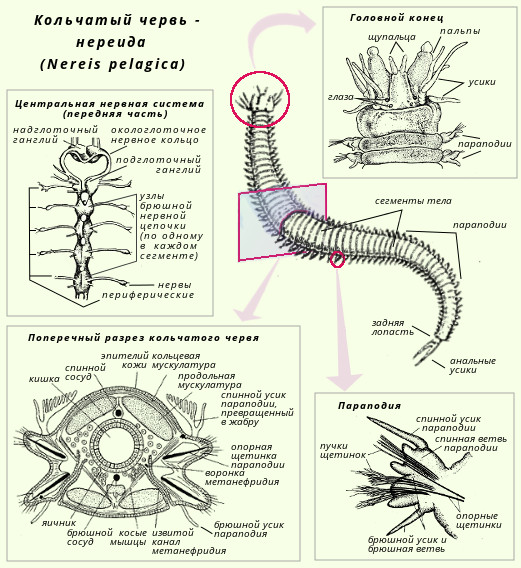
Целомические мешки в каждом сегменте помещаются между кишечником и стенкой тела, они заполнены водянистой жидкостью, в которой плавают амебоидные клетки.
Целом выполняет опорную функцию. Кроме того, в целомическую жидкость из кишечника поступают питательные вещества, которые затем распределяются по всему организму. В целоме накапливаются вредные продукты обмена веществ, которые удаляются выделительными органами. В стенках целома развиваются мужские и женские половые железы.
Центральная нервная система представлена надглоточным нервным узлом и брюшной нервной цепочкой. К надглоточному узлу проходят нервы от органов чувств: глаз, органов равновесия, щупалец и пальп. Брюшная нервная цепочка состоит из узлов (по одной паре в каждом сегменте тела) и стволов, связывающих узлы между собой. Каждый узел иннервирует все органы данного сегмента.
Пищеварительная система состоит из передней, средней и задней кишок. Передняя кишка обычно разделена на ряд отделов: глотку, пищевод, зоб и мускульный желудок. Рот находится на брюшной стороне первого сегмента тела. Задняя кишка открывается заднепроходным отверстием на задней лопасти. В стенке кишечника имеется мускулатура, обеспечивающая продвижение пищи.
Органы выделения - метанефридии - представляют собой парные трубчатые органы, метамерно повторяющиеся в сегментах тела. В отличие от протонефридиев они имеют сквозной выделительный каналец. Последний начинается воронкой, открывающейся в полость тела. Через воронку в нефридий поступает полостная жидкость. От воронки отходит каналец нефридия, открывающийся порой наружу. Проходя по канальцу, жидкость изменяет свой состав; в ней концентрируются конечные продукты диссимиляции, которые через наружную пору нефридия выбрасываются из организма.
У аннелид впервые в филогенезе животного мира появляется кровеносная система. Главные кровеносные сосуды проходят по спинной и брюшной стороне. В передних сегментах они соединены поперечными сосудами. Спинной и передние кольцевые сосуды способны ритмично сокращаться и выполняют функцию сердца. У большинства видов кровеносная система замкнутая: кровь циркулирует по системе сосудов, нигде не прерываемых полостями, лакунами или синусами. У некоторых видов кровь бесцветная, у других красная в связи с присутствием гемоглобина.
Большинство видов кольчатых червей дышит через кожу, богатую кровеносными капиллярами. У ряда морских форм имеются специализированные органы дыхания - жабры. Обычно они развиваются на параподиях или на пальпах. К жабрам подходят сосуды, несущие венозную кровь; она насыщается кислородом и в виде артериальной крови поступает в тело червя. Среди аннелид есть раздельнополые и гермафродитные виды. Половые железы расположены в полости тела.
Кольчатые черви обладают самой высокой организацией по сравнению с другими типами червей (см. табл. 1); у них впервые появляются вторичная полость тела, кровеносная система, органы дыхания, более высоко организована нервная система.
| Таблица 1. Характерные особенности различных типов червей | ||||||
| Тип | Кожно-мускульный мешок | Пищеварительная система | Кровеносная система | Половая система | Нервная система | Полость тела |
| Плоские черви | Включает слои продольных и кольцевых мышц, а также пучки спинно-брюшных и диагональных мышц | Из эктодермальной передней кишки и энтодермальной средней кишки | Не развита | Гермафродитная | Парный мозговой узел и несколько пар нервных стволов | Отсутствует, заполнена паренхимой |
| Круглые черви | Только продольные мышцы | Из эктодермальной передней и задней кйшки и энтодермальной средней кишки | То же | Раздельнополая | Окологлоточное нервное кольцо и 6 продольных стволов | Первичная |
| Из наружных кольцевых и внутренних продольных мышц | Из эктодермальной передней и задней кишки и энтодермальной средней кишки | Хорошо развита, замкнутая | Раздельнополые или гермафродиты | Парный мозговой узел, окологлоточное нервное кольцо, брюшная нервная цепочка | Вторичная | |
|
Для животных, относящихся к типу кольчатых червей, или кольчецов, характерны:
Кольчатые черви обитают в пресных и морских водах, а также в почве. Несколько видов обитает в воздушной среде. Главнейшими классами типа кольчатых червей являются:
Класс многощетинковые кольчецыС точки зрения филогенеза животного мира полихеты - наиболее важная группа аннелид, так как с их прогрессивным развитием связано возникновение вышестоящих групп беспозвоночных. Тело полихет сегментировано. Имеются параподии, состоящие из спинной и брюшной ветвей, каждая из которых несет на себе усик. В мускульной стенке параподиев имеются толстые опорные щетинки, а из вершины обоих ветвей торчат пучки тонких щетинок. Функция параподий различна. Обычно это локомоторные органы, участвующие в движении червя. Иногда спинной усик разрастается и превращается в жабру. Кровеносная система у полихет хорошо развита и всегда замкнутая. Имеются виды с кожным и жаберным дыханием. Полихеты - раздельнополые черви. Обитают в морях, преимущественно в прибрежной зоне. Характерным представителем класса может служить нереида (Nereis pelagica). Она в изобилии встречается в морях нашей страны; ведет донный образ жизни, будучи хищником, захватывает добычу челюстями. Другой представитель - пескожил (Arenicola marina) - обитает в морях, роет норы. Питается, пропуская через свой пищеварительный тракт морской ил. Дышит жабрами. Класс малощетинковые кольчецыМалощетинковые ведут свое происхождение от полихет. Наружными придатками тела служат щетинки, которые сидят непосредственно в стенке тела; параподий нет. Кровеносная система замкнутая; дыхание кожное. Малощетинковые кольчецы - гермафродиты. Подавляющее большинство видов - обитатели пресных вод и почвы. Характерным представителем класса может служить дождевой червь (Lumbricus terrestris). Дождевые черви обитают в почве; днем они сидят в норах, а вечером часто выползают наружу. Роясь в почве, они пропускают ее через свой кишечник и питаются содержащимися в ней растительными остатками. Дождевые черви играют большую роль в почвообразовательных процессах; они взрыхляют почву и способствуют ее аэрации; затаскивают в норы листья, обогащая почву органическими веществами; извлекают глубокие слои почвы на поверхность, а поверхностные - увлекают вглубь. Строение и размножение дождевого червя Дождевой червь имеет почти круглое в поперечном разрезе тело длиной до 30 см; насчитывают 100-180 сегментов или члеников. В передней трети тела дождевого червя находится утолщение - поясок (его клетки функционируют в период полового размножения и откладки яиц). По бокам каждого сегмента развиты по две пары коротких упругих щетинок, которые помогают животному при передвижении в почве. Тело имеет красновато-коричневую окраску, светлее на плоской брюшной и темнее на выпуклой спинной стороне. Характерной особенностью внутреннего строения является то, что у дождевых червей развиты настоящие ткани. Снаружи тело покрыто слоем эктодермы, клетки которой образуют покровную ткань. Кожный эпителий богат слизистыми железистыми клетками. Под кожей находится хорошо развитая мускулатура, состоящая из слоя кольцевых и находящегося под ним более мощного слоя продольных мышц. При сокращении кольцевых мышц тело животного вытягивается и становится тоньше, при сокращении продольных оно утолщается и раздвигает частички почвы.
Пищеварительная система начинается на переднем конце тела ротовым отверстием, из него пища поступает последовательно в глотку, пищевод (у дождевых червей в него впадают три пары известковых желез, поступающая из них в пищевод известь служит для нейтрализации кислот гниющих листьев, которыми питаются животные). Затем пища переходит в расширенный зоб, и небольшой мускульный желудок (мышцы в его стенках способствуют перетиранию пищи). От желудка почти до заднего конца тела тянется средняя кишка, в которой под действием ферментов пища переваривается и всасывается. Непереваренные остатки поступают в короткую заднюю кишку и выбрасываются наружу через заднепроходное отверстие. Дождевые черви питаются полусгнившими остатками растений, которые они глотают вместе с землей. При прохождении по кишечнику почва хорошо перемешивается с органическими веществами. Экскременты дождевых червей содержат в пять раз больше азота, в семь раз больше фосфора и в одиннадцать раз больше калия, чем обычная почва. Кровеносная система замкнутая, состоит из кровеносных сосудов. Вдоль всего тела над кишечником тянется спинной сосуд, а под ним - брюшной. В каждом сегменте их объединяет кольцевой сосуд. В передних сегментах некоторые кольцевые сосуды утолщены, их стенки сокращаются и ритмически пульсируют, благодаря этому кровь перегоняется из спинного сосуда в брюшной. Красный цвет крови обусловлен наличием в плазме гемоглобина. Для большинства кольчатых червей, в том числе и дождевых, характерно кожное дыхание, практически весь газообмен обеспечивается поверхностью тела, поэтому дождевые черви очень чувствительны к влажности почвы и не встречаются в сухих песчаных почвах, где их кожа скоро подсыхает, а после дождей, когда в почве много воды, выползают на поверхность. Выделительная система представлена метанефридиями. Метанефридий начинается в полости тела воронкой (нефростом) от которой идет проток - тоненькая петлеобразно изогнутая трубочка, открывающаяся наружу выделительной порой в боковой стенке тела. В каждом сегменте червя пара метанефридиев - правый и левый. Воронка и проток снабжены ресничками, вызывающими движение экскреторной жидкости. Нервная система имеет типичное для кольчатых червей строение (см. табл. 1), два брюшных нервных ствола, их узлы соединены между собой и образуют брюшную нервную цепочку. Органы чувств развиты очень слабо. У дождевого червя нет настоящих органов зрения, их роль выполняют отдельные светочувствительные клетки, находящиеся в кожном покрове. Там же помещаются рецепторы осязания, вкуса, обоняния. Как и гидра, дождевые черви способны к регенерации. Размножение происходит только половым путем. Дождевые черви - гермафродиты. В передней части их тела находятся семенники и яичники. Оплодотворение дождевых червей перекрестное. Во время копуляции и откладки яиц клетки пояска на 32-37-м сегменте выделяют слизь, служащую для образования яйиевого кокона, и белковую жидкость для питания развивающегося зародыша. Выделения пояска образуют своеобразную слизистую муфту. Червь выползает из нее задним концом вперед, откладывая в слизь яйца. Края муфты слипаются и образуется кокон, который остается в земляной норке. Эмбриональное развитие яиц происходит в коконе, из него выходят молодые черви. Ходы дождевых червей находятся преимущественно в поверхностном слое почвы до глубины 1 м, на зиму они спускаются до глубины 2 м. Через норки и ходы червей в почву проникают атмосферный воздух и вода, необходимые для корней растений и жизнедеятельности почвенных микроорганизмов. Через свой кишечник за сутки червь пропускает столько почвы, сколько весит его тело (в среднем 4-5 г). На каждом гектаре земли дождевые черви ежесуточно перерабатывают в среднем 0,25 т почвы, а за год они выбрасывают на поверхность в виде экскрементов от 10 до 30 т переработанной ими почвы. В Японии разводят специально выведенные породы быстро размножающихся дождевых червей и используют их экскременты для биологического метода обработки почвы. В выращиваемых на такой почве овощах и фруктах повышается содержание сахара. На важную роль дождевых червей в процессах почвообразования впервые указал Ч. Дарвин. Кольчатые черви играют значительную роль в питании донных рыб, так как местами черви составляют до 50-60 % биомассы придонных слоев водоемов. В 1939-1940 гг. из Азовского в Каспийское море был переселен червь нереис, составляющий ныне основу пищевого рациона осетровых рыб Каспийского моря.
Класс пиявкиТело сегментировано. Кроме истинной метамерии, имеется ложная кольчатость - по несколько колец в одном сегменте. Параподий и щетинок нет. Вторичная полость тела редуцировалась; вместо нее имеются синусы и щели между органами. Кровеносная система незамкнутая; кровь лишь часть своего пути проходит по сосудам и изливается из них в синусы и лакуны. Органов дыхания нет. Половая система гермафродитная. Медицинские пиявки специально разводятся и затем рассылаются в больницы. Используются, например, при лечении глазных болезней, связанных с увеличением внутриглазного давления (глаукома), при кровоизлиянии в мозг и гипертонии. При тромбозах и тромбофлебитах гирудин понижает свертываемость крови и способствует растворению тромбов. |
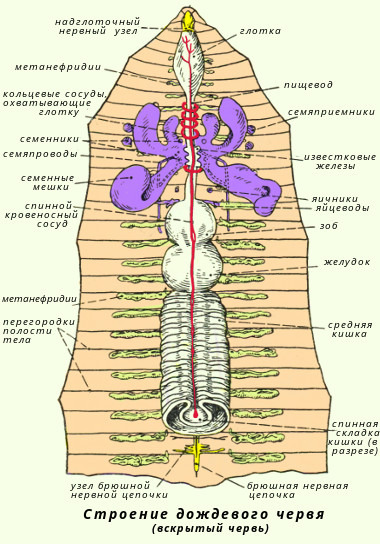
Кольчатые черви – беспозвоночные животные, среди которых ученые выделяют примерно 12 тысяч видов малощетинковых, многощетинковых червей, мизостомид и пиявок.
Описание кольчатых червей
Длина тела разных видов кольчатых червей варьируются от нескольких миллиметров до 6-ти метров. Тело кольчатого червя имеет билатеральную симметрию. Оно разделяется на область хвоста, головы и среднюю часть, которые состоят из многочисленных повторяющихся сегментов. Все сегменты тела отделены перегородками. В каждом из них содержится полный набор органов.
Рот находится в первом сегменте. Тело кольчатого червя заполнено жидкостью, благодаря которой образуется гидростатическое давление и придается форма телу. Наружный слой образован двумя слоями мышц. Волокна одного слоя располагаются в продольном направлении, а во втором слое они работают по круговой схеме. Передвижение осуществляется благодаря действию мышц, расположенных по всему телу.
Мышцы кольчатых червей могут работать таким образом, что части тела могут то удлиняться, то становится толстыми.
Образ жизни кольчатых червей
Кольчатые черви распространены по всему миру. В основном они обитают в земле и воде, но некоторые виды кольчатых червей кровососущие. Среди кольчатых червей имеются хищники, фильтраторы и падальщики. Самое большое экологическое значение имеют аннелиды, которые перерабатывают почву. К аннелидам относятся не только малощетинковые черви, но и пиявки. На 1 квадратный метр почвы может приходиться 50-500 червей.
Наиболее многообразны морские формы кольчатых червей. Они живут во всех широтах Мирового океана и могут встречаться на разных глубинах, вплоть до 10 километров. Они имеют высокую плотность поселения: на 1 квадратный метр приходится около 500-600 морских кольчатых червей. Кольчатые черви очень важны в морской экосистеме.
 Кольчатые черви — раздельнополые животные, некоторые являются гермафродитами.
Кольчатые черви — раздельнополые животные, некоторые являются гермафродитами. Размножение кольчатых червей
Многие виды кольчатых червей размножаются бесполым путем, но имеются виды, размножающиеся половым путем. Большая часть видов развивается из личинок.
Для полихет и олигохет характерна способность к регенерации, поэтому они размножаются вегетативным способом. У некоторых видов, к примеру, у аулофорусов, при наличии достаточного количества пищи, на сегментах тела образуются дополнительные ротовые отверстия, по которым со временем происходит отделение и образование новых особей – дочерних клонов.
Питание кольчатых червей

Классификация кольчатых червей
Кольчатых червей считают близкими сородичами членистоногих. Они имеют общие черты: сегментированное тело и строение нервной системы. Самое большое сходство с членистоногими имеют многощетинковые черви. У них также развиты боковые придатки – параподии, которые считаются зачатками ног.
По типу дробления и строению личинок кольчатые черви имеют сходство с моллюсками и сипункулидами.
Считается, что ближайшими сородичами кольчатых червей являются плеченогие, немертины и форониды, моллюски — более дальние родственники, а самые дальние сородичи – плоские черви.
В разных классификациях выделяют разное количество классов кольчатых червей. Но традиционно их подразделяют на 3 класса: малощетинковые, многощетинковые и пиявки. Также существует другая систематика:
Многощетинковые черви – этот класс является самым многочисленным, и он состоит преимущественно из морских форм;
Мизостомиды;
Поясковые черви с характерным пояском на теле.
Эволюция кольчатых червей
Имеется несколько версий о происхождении кольчатых червей. Как правило, считается, что они произошли от низших плоских червей. Определенные черты свидетельствуют, что кольчатые черви имеют общее сходство с низшими червями.

Предполагается, что первыми произошли многощетинковые черви, а от них образовались пресноводные и наземные формы – малощетинковые черви и пиявки.
Найдено немного ископаемых останков кольчатых червей с периода среднего кембрия. К кембрийским ископаемым относится вид Canadia. У этих червей не имелось челюстей, но позже у некоторых многощетинковых сформировался челюстной хитиновый аппарат, о чем свидетельствуют найденные останки сколекодонты. Челюстной аппарат имеет вид искривленных, зазубренных пластид длиной 0,3-3 миллиметра.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .

Тело кольчецов расчленено на головной отдел (простомиум ), следующие за ним кольца (или сегменты, или метамеры ), число которых, как правило, велико (несколько десятков), и задний отдел (анальную лопасть, или пигидиум ). Головной отдел у морских червей, называемых многощетинковыми, хорошо выражен и несет на себе разные придатки: широкие, узкие и т. д. (рис. 61). У пресноводных и наземных кольчецов головной отдел выражен слабо (рис. 61). С простомиумом может срастаться несколько передних колец. Сегменты тела, как правило, сходны по своему строению. Такое расчленение называется гомономной сегментацией или гомономией метамерией. Оно не только наружное, но глубоко внутреннее, так как каждый сегмент отделен от соседних перегородками и имеет комплект органов.
Кожный покров состоит из однослойного эпителия и выделенной им тонкой кутикулы (рис. 62). В коже много желез, которые выделяют слизь, облегчающую передвижение червей, и другие секреты (например, вещества, способствующие у раздельнополых кольчецов привлечению самок к самцам, ядовитые для других животных, и т. д.).
Нервная система. Эта система развита значительно лучше, чем у других червей, и в ее строении очень ярко отражено расчленение тела кольчецов на сегменты. Центральный отдел ее состоит, как правило, из двух головных узлов, лежащих на спинной стороне, окологлоточных тяжей, переходящих на брюшной стороне в цепочку, обычно очень длинную и образующую в каждом сегменте узел (рис. 63, Б), чем объясняется ее название. Таким образом, брюшная цепочка образовалась из двух тяжей. У низших форм типа тяжи остаются разделенными на всем своем протяжении и соединены перемычками, что напоминает лестницу (рис. 63, А). Такая система менее централизована, она похожа на центральную нервную систему низших червей - плоских и первичнополостных (см. рис. 31, Б, и 54).
Узлы и тяжи типичных кольчатых червей развиты значительно лучше и строение их более сложно, чем у последних. Вся центральная система кольчецов отделена от эпидермиса, в то время как у низших червей она еще соединена с эпидермисом. Каждый узел брюшной цепочки иннервирует и воздействует на работу органов, расположенных в кольце, где находится узел. Головные узлы, развитые лучше, чем узлы цепочки, координируют работу последних и через них деятельность всего тела. Кроме того, они иннервируют глаза и другие органы чувств, расположенные в головном отделе тела.
Органы чувств разнообразны. В коже рассеяны осязательные клетки, которых особенно много на придатках тела. Имеются органы, воспринимающие химические раздражения. Светочувствительные органы имеются у всех кольчатых червей. Самые простые из них представлены специальными клетками, рассеянными по всей коже. Поэтому почти у всех кольчецов кожа чувствительна к световым раздражениям. На переднем конце тела, а у ряда пиявок и на заднем, светочувствительные органы усложняются и превращаются в глаза. У ряда форм имеются органы равновесия, похожие по своему строению на аналогичные органы медуз и других низших животных.
Прогрессивное развитие нервной системы кольчатых червей обеспечивает более сложные и энергичные движения их тела, активную работу всех систем органов, лучшее согласование функций всех частей организма, более сложное поведение и делает возможным более тонкое приспособление этих животных в окружающей среде.
Двигательная система. Эта система у кольчатых червей более совершенная, чем у ранее изученных червей. Ресничное движение свойственно только личинкам, у взрослых форм, за редким исключением, оно отсутствует, и их передвижение совершается только благодаря работе мышц. Кожно-мускульный мешок развит значительно лучше, чем у плоских и первичнополостных червей (ср. рис. 32, 53 и 62). Под эпидермисом лежит хорошо развитый слой кольцевых мышц (рис. 62), состоящих из длинных волокон с ядрами. При сокращении этих мышц тело червя становится более тонким и длинным. За кольцевыми мышцами идет значительно более толстый слой продольных мышц, сокращение которых укорачивает тело и делает его более толстым. Одностороннее сокращение продольных и некоторых других мышц приводит к изгибу тела и к изменению направления движения. Кроме того, имеются мышцы, идущие от спинной стороны к брюшной: мышцы, проходящие в перегородках, разделяющих кольца; мышцы различных придатков тела, играющих вспомогательную роль в передвижении червей, и др. Сила мышц кожно-мускульного мешка велика и позволяет червям бысгро проникать глубоко в грунт. Многие кольчатые черви могут плавать. Опорой для мышц в основном служит гидроскелет, образуемый жидкостью полости тела, а также пограничные образования.
Движение кольчатых червей облегчается вспомогательными придатками (см. рис. 61, 62, 64): щетинками (имеются у подавляющего большинства видов) и параподиями (имеются у большинства морских червей). Щетинки (см. рис. 62, 64, А, Б) представляют собой твердые образования из органического вещества, очень сложного углевода - хитина, разной формы, толщины и длины. Щетинки образуются и приводятся в движение специальными мышечными пучками. Щетинки расположены (одиночно или пучками) правильными продольными рядами почти на всех кольцах червей. Параподии (рис. 64,В) представляют собой мощные боковые выросты тела с хорошо развитой мускулатурой. Параподии соединены с телом подвижно, и эти придатки действуют по типу простого рычага. Каждая параподия обычно состоит из двух лопастей: спинной и брюшной, которые, в свою очередь, могут подразделяться на лопасти второго порядка. Внутри каждой из главных лопастей имеется опорная щетинка. Параподии несут пучки щетинок, далеко выдающихся за пределы тела. На параподии имеются два щупика - спинной и брюшной, в эпидермисе которых находятся различные органы чувств, воспринимающие механические и другие раздражения. Передвижение кольчатых червей сильно облегчается расчленением их на кольца, в результате чего увеличивается гибкость тела.
В теле кольчецов имеются уплотненные пластинки, называемые пограничными образованиями , которые подстилают эпидермис, разделяют мышцы, сильно развиты в перегородках между кольцами. Они придают прочность всему телу, служат опорой для двигательного аппарата, важны для функционирования кровеносной и пищеварительной систем и выполняют защитную роль.
Кровеносная система. У кольчатых червей в связи со значительным усложнением строения их тела и резко возросшей активностью их жизнедеятельности развилась более совершенная система переноса веществ - кровеносная. В ее состав входят два главных сосуда - спинной и брюшной (рис. 62 и 65). Первый проходит над кишкой, вплотную подходя к его стенкам, второй - под кишкой. В каждом сегменте оба сосуда соединены кольцевыми сосудами. Кроме того, имеются более мелкие сосуды - особенно много их в стенках кишечника, в мускулатуре, в коже (через которую происходит обмен газов), в перегородках, разделяющих сегменты тела, и т. д. Кровь движется благодаря сокращению самих сосудов, главным образом спинного и передних кольцевых, в стенках которых хорошо развиты мышечные элементы.
Кровь состоит из жидкой части - плазмы , в которой плавают кровяные клетки - форменные элементы крови . В плазме имеются дыхательные пигменты, т. е. особые сложные органические соединения. Они поглощают кислород в органах дыхания и отдают его тканям организма. У некоторых кольчецов в плазме имеется один из наиболее совершенных дыхательных пигментов - гемоглобин; у этих кольчецов цвет крови красноватый. Большей частью в крови кольчатых червей находятся другие пигменты и цвет ее бывает зеленоватый, желтоватый и т. д. Клетки крови довольно разнообразны. Среди них есть фагоциты, выпускающие, подобно амебам, ложноножки, захватывающие бактерии, всякие инородные тела, отмирающие клетки тела и переваривающие их. Как отмечалось ранее, фагоциты имеются у всех животных. Таким образом, кровеносная система не только обеспечивает перенос разных веществ, но выполняет и другие функции.
Полость тела. Полость тела кольчецов отличается по строению от первичной полости. Последняя не имеет собственных стенок: с наружной стороны она ограничивается мышцами кожно-мускульного мешка, с внутренней - стенкой кишечника (см. рис. 53). Полость же тела кольчатых червей, называемая вторичной или целомом , окружена однослойным эпителием, который, с одной стороны, прилегает к кожно-мускульному мешку, а с другой - к кишечнику (см. рис. 62). Следовательно, стенка кишечника становится двойной . Целом наполнен водянистой жидкостью, постоянно находящейся в движении, в которой плавают клетки, похожие на клетки крови (фагоциты, клетки с дыхательными пигментами и др.). Таким образом, вторичная полость тела кроме роли гидроскелета выполняет функции, сходные с таковыми крови (перенос веществ, защита от болезнетворных организмов и др.). Однако следует подчеркнуть, что целомическая жидкость движется медленнее крови и она не может войти в такое тесное соприкосновение со всеми частями тела, как разветвленная сеть капилляров.
Дыхательная система. У кольчатых червей обмен газов в основном происходит через кожу, но процессы дыхания в связи с появлением кровеносной системы и целома у них более совершенны, чем у ранее рассмотренных червей. Многие кольчецы, преимущественно морские, имеют разветвленные придатки, играющие роль жабр (см. рис. 61, Б). Дыхательная поверхность увеличивается также благодаря наличию разных выростов тела. Усовершенствование процессов дыхания имеет большое значение для кольчатых червей в связи с активизацией их образа жизни.
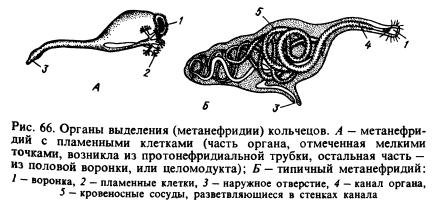
Выделительная система. Основными органами выделения являются метанефридии (рис. 66, Б). Типичный метанефридий состоит из воронки и длинной извитой трубочки, в стенках которой разветвляются кровеносные сосуды. В каждом сегменте, за исключением некоторых, по два этих органа, слева и справа от кишечника (см. рис. 65). Воронка обращена в полость одного сегмента, а трубочка пронизывает перегородку, проходит в другой сегмент и открывается наружу на брюшной стороне тела. Продукты диссимиляции извлекаются метанефридиями из целомической жидкости и из оплетающих их кровеносных сосудов.
У ряда кольчатых червей с метанефридиями связаны трубочки протонефридиального типа, замкнутые на концах, обращенных в полость тела, пламенными клетками. Возможно, что метанефридии возникли из протонефридий, которые соединились с воронками, развившимися на перегородках между кольцами (рис. 66, А). Предполагают, что эти воронки, называемые целомодуктами , первоначально служили для выхода из полости тела половых продуктов.
На стенках целома имеются многочисленные клетки, поглощающие из полостной жидкости продукты распада. Особенно много таких клеток, называемых хлорагогенными , имеется на стенках средней части кишечника. Продукты распада, изъятые из целомической жидкости и заключенные в названных клетках, не могут больше оказывать вредное воздействие на организм. Клетки, нагруженные такими продуктами, могут через метанефридии или через поры в стенках тела выходить наружу.
Пищеварительная система. Пищеварительная система кольчецов (см. рис. 65) в связи с более активным, чем у ранее рассмотренных групп животных, образом жизни и прогрессом всей организации тоже более совершенна. У кольчецов: 1) больше выражено разделение пищеварительной системы на различные отделы, каждый из которых выполняет свою функцию; 2) строение стенок пищеварительной трубки более сложное (более развиты пищеварительные железы, мускулатура и др.), в результате чего лучше обрабатывается пища; 3) кишечник связан с кровеносной системой, благодаря чему переваривание пищевых веществ и их всасывание идет более интенсивно и улучшено снабжение его веществами, необходимыми для выполняемой им работы.
Пищеварительная трубка, как правило, прямая и разделяется на следующие отделы: ротовая полость, глотка, пищевод, который может расширяться в зоб, мускулистый желудок (имеется у ряда видов, например у дождевых червей), средняя кишка (обычно очень длинная), задняя кишка (сравнительно короткая), открывающаяся наружу анальным отверстием. В глотку и пищевод впадают протоки желез, секрет которых имеет значение в обработке пищи. У многих хищных многощетинковых кольчецов глотка вооружена челюстями, передняя часть пищеварительной трубки может выворачиваться в виде хобота, что помогает овладеть жертвой и проникнуть в ее тело. Средняя кишка у ряда видов имеет глубокое впячивание (тифлозоль ), тянущееся вдоль всей спинной стороны этой кишки (см. рис. 62). Тифлозоль увеличивает поверхность кишечника, что ускоряет переваривание и всасывание пищи.
Размножение. Одни кольчецы размножаются бесполым и половым способами, у других наблюдается только половое размножение. Бесполое размножение происходит путем деления. Часто в результате деления может получиться цепочка червей, которые не успели еще разойтись.
Строение полового аппарата различно. Многощетинковые кольчецы (они обитают в морях) раздельнополы и имеют просто устроенный половой аппарат. Половые железы развиваются у них на стенках целома, половые клетки выходят в воду через разрывы в стенках тела или через метанефридии и оплодотворение яйцеклеток происходит в воде. Кольчецы, живущие в пресной воде и в сырой земле (малощетинковые), а также все пиявки гермафродиты, их половой аппарат имеет сложное строение, оплодотворение внутреннее.
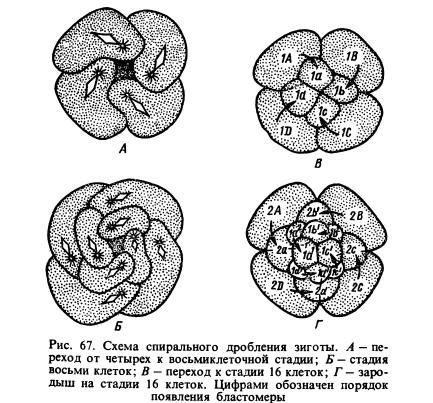
Развитие. Дробление оплодотворенного яйца, в результате которого образующиеся бластомеры располагаются по спирали (рис. 67), напоминает те же процессы у ресничных червей. Многощетинковые кольчецы развиваются с превращением: из их яиц образуются личинки трохофоры (рис. 68), совсем не похожие на взрослых червей и превращающиеся в последних только после сложных преобразований. Трохофора - планктонный организм. Она очень мала, прозрачна, по экватору ее тела обычно проходят два пояса ресничек: один, верхний, над ртом, другой, нижний, под ртом. Следовательно, трохофора состоит из двух частей: верхней, или передней, и нижней, или задней, заканчивающейся анальной лопастью. У трохофор некоторых видов может быть несколько поясов ресничек. На верхнем конце торчит пучок ресничек, прикрепленных к темянной пластинке (личиночному органу чувств). Под пластинкой находится нервный центр, от которого отходят нервы. Мышечная система состоит из волокон, идущих в разных направлениях. Кровеносной системы нет. Пространство между стенками тела и кишечником представляет собой первичную полость тела. Органы выделения - протонефридии. Пищеварительный аппарат состоит из трех отделов: переднего, среднего и заднего, заканчивающегося анальным отверстием. Благодаря работе ресничек личинка движется и в рот поступает пища, состоящая из микроскопических организмов и органических кусочков. Некоторые трохофоры активно захватывают ртом мелких животных. По своему строению трохофора напоминает первичнополостных червей, но в некоторых отношениях она похожа и на личинок морских ресничных червей. Стенки тела, нервная система, протонефридии, начало и конец пищеварительного аппарата трохофоры образовались из эктодермы, больщая часть кишечника - из энтодермы, мышечные волокна - из клеток, называемых мезенхимными и происходящих из обоих слоев.
При превращении трохофоры во взрослого червя она претерпевает ряд существенных изменений. В этих изменениях важнейшую роль играют зачатки третьего зародышевого листка - мезодермы . Одни зачатки мезодермы имеются еще у личинки до начала метаморфоза, они лежат с каждой стороны между стенками тела и задней частью кишечника (рис. 68,В,12). Другие зачатки мезодермы образуются позднее от переднего края анальной лопасти, который превращается в зону роста червя (рис. 68, В, 13). Метаморфоз личинки начинается с того, что задняя часть ее удлиняется и перетяжками стенок тела подразделяется на 3, 7, редко больше сегментов. После этого удлиняются и зачатки мезодермы, лежащие между стенками тела и задней частью кишечника, и делятся на столько же участков, сколько образовалось сегментов в результате наружных перетяжек. В каждом кольце их два (рис. 68, Д, 14). Сегменты, образовавшиеся из задней части трохофоры, называются личиночными или ларвальными , они характерны для поздних стадий развития трохофоры, когда она уже начинает немного походить на взрослого червя, но еще имеет мало сегментов. В процессе дальнейшего развития сегменты образуются упомянутой выше зоной роста. Эти сегменты называются послеличиночными , или постларвальными (рис. 68, Д). Их образуется столько, сколько сегментов имеет взрослый червь данного вида. В постларвальных сегментах сначала делятся на участки мезодермальные зачатки (по два в каждом кольце), а потом наружные покровы.
Основные системы органов взрослого червя образуются следующим образом (рис. 69, А). Из эктодермы развиваются эпидермис, нервная система, передний и задний концы пищеварительной трубки. Мезодермальные зачатки в каждом кольце растут и вытесняют при этом первичную полость. В конце концов правые и левые зачатки сходятся над и под кишечником, так что вдоль него, сверху и снизу, образуются спинной и брюшной кровеносные сосуды. Следовательно, стенки сосудов образуются из мезодермы, а полость их представляет собой остатки первичной полости тела. В середине же зачатков клетки раздвигаются, возникает и все разрастается целомическая полость тела, которая со всех сторон окружена клетками мезодермального происхождения. Такой способ образования целома называется телобластическим . Каждый мезодермальный зачаток, разрастаясь, сходится впереди и сзади с соседними зачатками (рис. 69, Б) и между ними возникают перегородки, а мезодермальные клетки, окружающие остатки первичной полости между перегородками, образуют кольцевые кровеносные сосуды. Наружный листок мезодермальных зачатков, прилегающий к эктодерме, дает начало мышцам, внутренний листок окружает пищеварительную трубку. Следовательно, стенки кишечника становятся теперь двойными: внутренний слой (за исключением переднего и заднего концов, происходящих из эктодермы) развился из энтодермы, наружный - из мезодермы. Воронки метанефридиев образуются из клеток мезодермального слоя, а трубки их (представляющие остатки протонефридиев) - из эктодермы.
Постепенно происходит развитие всех частей тела взрослого червя; дифференцируются слои мышц, увеличивается количество кровеносных сосудов, кишечник разделяется на отделы, в его стенках развиваются железистые клетки, мышечные волокна, кровеносные сосуды и т. д. Головная лопасть (простомиум) взрослого червя образуется из верхней части трохофоры, кольца тела из ларвальных и постларвальных сегментов, а пигидиум - из анальной лопасти личинки.
Происхождение . О происхождении кольчатых червей высказаны разные гипотезы. Сторонники одной гипотезы полагают, что кольчатые черви произошли от турбеллярий. Действительно, в эмбриональном развитии обеих групп животных есть сходные черты. Центральная нервная система кольчецов (т. е. головные узлы и брюшная цепочка) могла образоваться из той же системы более сложных турбеллярий, у которых узлы переместились в передний конец тела и из продольных тяжей осталось два главных и таким образом возникла центральная нервная система лестничного типа, сохранившаяся и у низших кольчатых червей. Кожно-мускульный мешок плоских червей мог превратиться в аналогичную систему кольчецов, а метанефридии могли возникнуть из протонефридий. Однако с эволюционной точки зрения невозможно допустить, что наиболее высокоорганизованные черви произошли непосредственно от самых низших червей, у которых нервная и мышечная системы были еще слабо развиты, нет полости тела, кишечник не дифференцирован еще на три отдела и пищеварение в основном остается внутриклеточным и т. д. Очевидно, предками высших червей были черви с более сложным строением, чем турбеллярии.
Согласно другой гипотезе начало кольчецам дали немертины, т. е. черви, несомненно, произошедшие от турбеллярий, но имевшие значительно более сложное строение, чем последние (значительное развитие нервной и мышечной системы, появление кровеносной системы, сквозного кишечника и др.). Автор этой гипотезы выдающийся советский зоолог Н. А. Ливанов высказал предположение, что у наиболее прогрессивной группы немертин в кожно-мускульном мешке возникли метамерно расположенные полости, служившие опорой для мышц и превратившиеся потом в целомические полости, в результате чего резко улучшилось движение животных. Противники этой гипотезы считают, что немертины, у которых одной из главных особенностей является хобот, отсутствующий у кольчецов, не могли быть предками последних. Однако нужно полагать, что хобот развился у немертин после длительной эволюции, когда у них появились более сильные, чем раньше, соперники в охоте на животных. Кольчатые же черви могли произойти от неспециализированных немертин, организация которых была уже сложна, но хобот не был развит. Другое возражение против рассматриваемой гипотезы более серьезно. Из этой гипотезы вытекает, что кровеносная система возникла раньше целома, а последний развился с самого начала в виде метамерных образований. Между тем известны черви, несомненно, родственные кольчатым червям, у которых метамерия еще не выражена, целом сплошной и нет кровеносной системы. Раньше полагали, что упомянутые черви упростились в связи с приспособлением к малоактивному образу жизни, но новые исследования подтверждают изначальную примитивность целомических червей, о которых идет речь.
Авторы третьей гипотезы считают, что предками кольчецов были первичнополостные черви, но не такие специализированные, как коловратки и круглые черви, а более близкие к предкам этого типа. Данная гипотеза основана главным образом на строении трохофоры, которая, как показано было выше, имеет важные черты сходства (первичная полость тела, протонефридии, сквозной кишечник) с первичнополостными червями, но еще лишена особенностей кольчатых червей. Приняв эту гипотезу, следует предположить, что целом возник в результате развития эпителия на стенках первичной полости тела, а метамерия тела и кровеносная система появились позднее. Из этой же гипотезы следует, что немертины, несмотря на прогрессивные черты своей организации, не имели отношения к появлению более высокоорганизованных типов животных. Наоборот, немертинная гипотеза происхождения кольчатых червей отвергает значение первичнополостных червей для формирования новых типов животных.
Рассматривать здесь достаточно обстоятельно разные возражения против каждой из упомянутых гипотез нельзя, так как для этого нужны более подробные сведения о строении и развитии всех типов червей, но не подлежит сомнению, что целомические черви не могли возникнуть непосредственно от самых низших червей.